
{kind=link}
{kind=link}
390份小麦-黑麦种质材料主要农艺性状分析及优异材料的GISH与FISH鉴定
[罗巧玲1, 4 , 郑琪2 , 许云峰1 , 李立会3 , 韩方普2 , 许红星1 , 李滨2 , 马朋涛1 , 安调过1, *  ]
]
]
|
|


将小麦近缘属植物黑麦中的优良基因导入小麦可以拓宽小麦的遗传基础, 丰富小麦的遗传变异。本研究调查并分析了390份小麦-黑麦种质材料。在这390份种质材料中, 6个主要农艺性状值均有较大的极差, 说明其遗传多样性丰富。与10份小麦主栽品种相比, 90%以上的材料具有穗长和分蘖数的显著优势, 60%以上的材料具有小穗数优势, 约30%的材料穗粒数和千粒重显著高于主栽品种。利用基因组原位杂交(genomic
Transferring desirable genes of rye (
小麦在世界上种植广泛, 是人类最重要的粮食作物之一。20世纪90年代以后, 由于缺少新的突破性育种亲本, 通过常规技术选育的品种(系)多出自相同或相似的亲本, 导致小麦的遗传多样性日趋降低[ 1]。从小麦近缘属中挖掘有益基因可以拓宽小麦的遗传基础, 丰富其遗传多样性[ 2, 3], 因而成为研究热点之一。
黑麦属中的栽培黑麦( Secale cereale L.)与小麦同属禾本科小麦族的小麦亚族(Tritticine), 其染色体组为RR (2 n = 2 x = 14)。栽培黑麦分蘖力强, 小穗数多, 抗多种病虫害, 根系发达, 具有抗旱、耐盐碱、耐贫瘠、耐寒和抗干热风等特性, 是小麦育种中改善重要性状和丰富遗传多样性的优良基因供体[ 4, 5, 6, 7]。小麦与黑麦远缘杂交得到的小黑麦是第一个人工创造的新物种, 也是黑麦染色体导入小麦形成的典型双二倍体[ 8]。小黑麦结合了小麦和黑麦的优良特性, 其本身就具有很高的生产价值。目前, 直接用于生产的主要是六倍体和八倍体小黑麦。另外, 与黑麦相比小黑麦更容易与普通小麦杂交成功, 所以它也常作为黑麦优良基因向小麦转移的桥梁亲本[ 9]。黑麦基因资源对小麦产量等性状的改良发挥了重要作用, 尤其是T1RS·1BL易位系在抗病、抗逆和产量等方面的突出表现使其在世界范围内得到广泛应用, 成为利用外源基因进行小麦遗传改良最成功的范例之一[ 10, 11, 12]。Robinovich等[ 11]调查发现, 全世界约330个小麦推广品种含有小麦-黑麦易位系。在我国, 具有黑麦或小黑麦血缘的小麦推广品种至少有30个[ 13]。
随着现代分子细胞遗传学和分子生物学的发展, 小麦中外源遗传物质的鉴定技术不断丰富和深化, 从传统的形态学标记、细胞学标记(染色体核型和染色体分带), 发展到生化标记(同工酶等)、原位杂交技术和目前应用广泛的分子标记等。其中, 荧光原位杂交是指将特殊修饰的核苷酸标记的DNA探针直接与染色体DNA杂交, 再用与荧光素分子偶联的单克隆抗体和探针分子间的特异性结合来检测DNA分子在染色体上位置、分布和相对含量的一项分子细胞遗传学技术[ 14]。它是架设在传统细胞遗传学和现代分子遗传学之间的“桥梁”。以全基因组DNA为探针与固定在玻片上的染色体进行基因组原位杂交(genomic in situ hybridization, GISH), 可在染色体水平上, 清晰地显示出远缘杂交后代中与探针同源的部分[ 15, 16]。如以黑麦基因组DNA为探针, 即可鉴定出导入小麦背景中的黑麦外源遗传物质。新发展起来的多色荧光原位杂交技术(multicolor fluorescent in situ hybridization, mc-FISH)是将特定的DNA重复序列标记为探针与染色体DNA杂交, 在不同的染色体上显示出不同的带型, 以此来确定染色体的归属[ 17, 18]。它已成为鉴定异源材料、识别染色体的新手段。顺序GISH与FISH技术不仅可以鉴定是否有外源黑麦遗传物质的导入, 还可以确定染色体的归属[ 19], 并且具有快速、直观、有效的特点, 对鉴定和跟踪导入的外源遗传物质, 加速种质创新、提高选择的准确性具有重要意义。
本研究以390份小麦-黑麦种质材料为研究对象, 调查其主要农艺性状, 并对其中具有代表性的8份材料进行了基因组原位杂交(GISH)和多色荧光原位杂交(mc-FISH)鉴定, 旨在明确优异材料及其染色体组成, 为这些材料在小麦育种中的进一步应用提供依据。
390份小麦-黑麦种质资源由中国农业科学院作物科学研究所的李立会研究员提供; 另外选择10个小麦主栽品种为对照, 分别是济麦22、石新828、科农199、良星99、石4185、石麦15、冀5265、山农21、烟农19和邯7086。
2012—2013小麦生长季, 将所有材料种植在中国科学院栾城实验站。每份材料种3行, 每行20粒, 行长1.5 m, 行距25 cm。试验区四周设保护行, 田间统一管理, 依据《小麦种质资源描述规范和数据标准》[ 20]考察农艺性状。小麦成熟后每小区随机选取中间行的中部植株5株, 分别调查株高和分蘖数, 取平均值; 取5个主穗, 调查穗长、小穗数、穗粒数和不育小穗数, 取平均值; 每小区收获中间行1 m长测产, 计算单位面积产量; 从收获株上取500粒称重, 计算千粒重, 3次重复, 取平均值。
利用SPSS 16.0软件进行单因素方差分析, 按LSD法比较小麦-黑麦种质材料与10个主栽品种的差异显著性。
分别选择分蘖数、小穗数、穗粒数和千粒重表现最好的4份材料, 产量较高的3份材料, 以及表型近似普通小麦的1份材料进行GISH和mc-FISH鉴定。
挑选饱满的种子在垫有双层湿滤纸的培养皿中23℃发芽, 露白后在4℃下作同步化处理24 h, 然后转入23℃恒温培养24 h, 剪取发芽种子的根。取细胞分裂中期的根尖, 按照韩方普描述的方法[ 21]进行染色体细胞制备、探针标记和原位杂交。GISH鉴定以中国春基因组DNA作为封阻, 黑麦基因组DNA作为探针, 按100∶1与制备的根尖中期细胞染色体原位杂交。Mc-FISH鉴定以来自 Aegilops squarrosa的重复序列p As1 (标记为红色)[ 22]和来自黑麦的重复序列p Sc119.2 (标记为绿色)[ 23]为探针, 与制备的根尖中期细胞染色体原位杂交; 小麦与黑麦染色体的标注参考已发表的图谱[ 17, 24]。
390份小麦-黑麦种质的分蘖数、穗长、小穗数、穗粒数和千粒重变异范围很大, 分蘖数相差27.8, 穗长相差9.0 cm, 小穗数相差19.2, 穗粒数相差74.0粒, 千粒重相差39.5 g (表1)。大部分材料的性状值集中在
| 表1 390份小麦-黑麦种质材料与10份小麦主栽品种主要农艺性状比较 Table 1 Comparison of 390 wheat-rye derivatives with 10 major wheat cultivars in main agronomic traits |
一定范围内, 如87.7%的材料分蘖数在9~21区间, 91.5%的材料穗长在9.5~15.5 cm区间, 83.3%的材料小穗数为18~27, 84.1%的材料穗粒数为41~65, 82.1%的材料千粒重集中在30.5~45.5 g范围内。此外, 390份材料株高变异范围为71.0~172.6 cm, 极差为101.6 cm, 其中67.4%的材料为100~130 cm, 还有7份材料株高超过150 cm, 仅14份材料(3.6%)的株高为70~80 cm (图1)。
超过90%的小麦-黑麦种质材料表现为分蘖数和穗长显著优于小麦主栽品种, 并且385份材料(98.7%)的分蘖数超过主栽品种的最小值7.0 (科农199); 所有材料的穗长都大于主栽品种的最小值7.2 cm (科农199); 小穗数也有明显优势, 其中254份材料(65.1%)显著优于主栽品种, 357份材料(91.5%)的小穗数高于主栽品种的最小值18.3 (山农21)。良星99、山农21和烟农19的不育小穗数都为0, 石新828、石麦15、冀5265和科农199都为1, 而260份(66.7%)小麦-黑麦种质材料的不育小穗数小于1 (表1)。穗粒数和千粒重两性状, 小麦-黑麦种质材料的优势相对较弱, 但也有43份材料(11.0%)的穗粒数大于主栽品种最大值65.0 (山农21), 204份材料(52.3%)大于主栽品种最小值52.4 (石新828); 38份材料(9.7%)的千粒重大于主栽品种的最大值43.3 g (良星99), 293份材料(75.1%)大于主栽品种最小值32.9 g (石4185)。
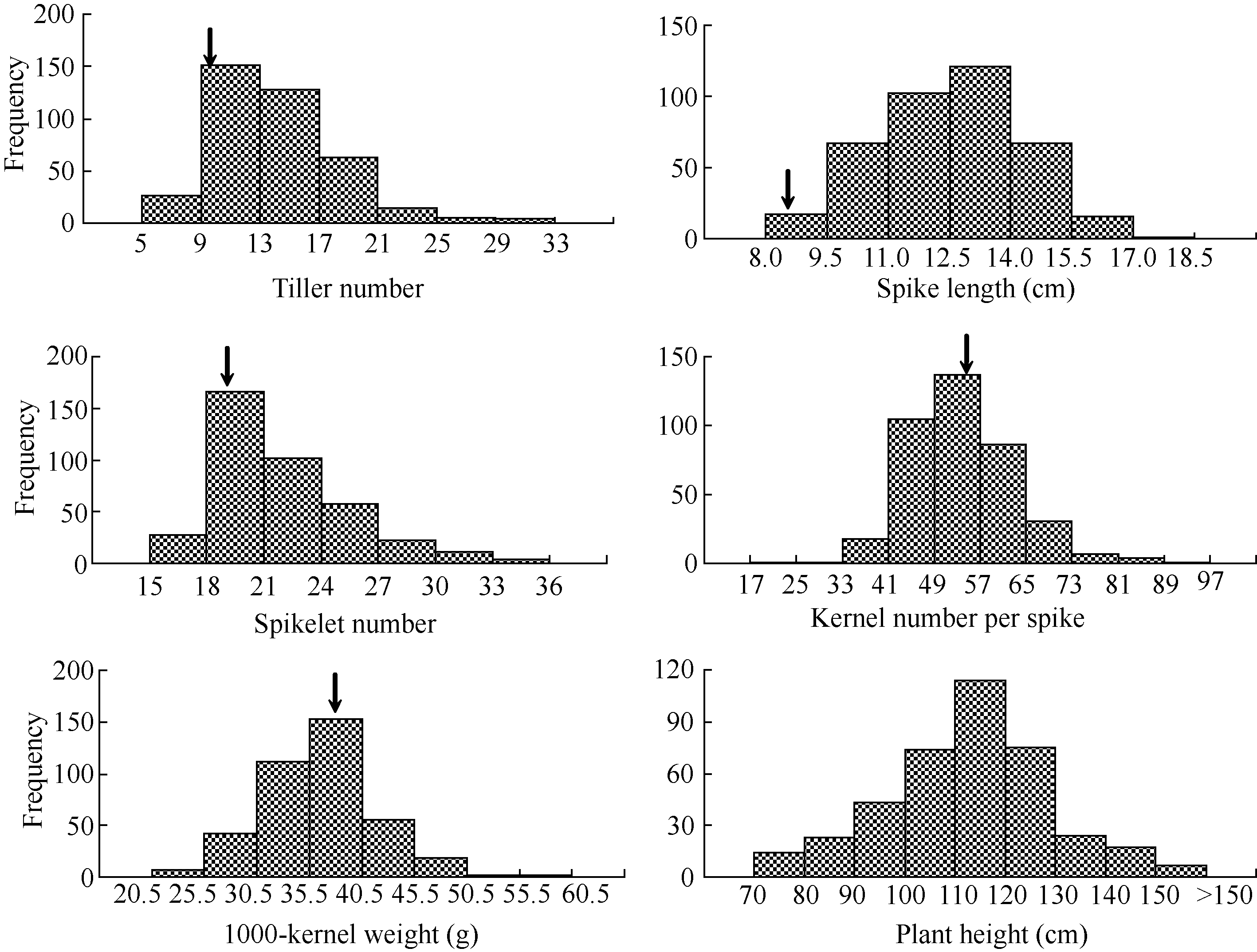 | 图1 390份小麦-黑麦种质6个农艺性状的频次分布箭头示10份主栽品种的平均值。Fig. 1 Distributions of frequency in six agronomic traits of 390 wheat-rye derivativesThe arrows show the means of 10 major wheat cultivars. |
| 表2 8份代表性材料的主要农艺性状 Table 2 Main agronomic traits of eight representative wheat-rye derivatives |
从390份小麦-黑麦种质中选出8份代表性材料, 其农艺性状(表2)与育种目标相符或在育种中有一定应用价值。其中R1162、R2203和R2221为高生物量和高产系, 均表现较大的穗长(15.9~16.2 cm)和较多的小穗(26.6~34.4), 产量三要素相对协调; R1147、R1103、R2209和R1017分别为390份材料中分蘖数最多、主穗小穗数最多、穗粒数最多和千粒重最大的材料, 虽然综合性状不如上述3份产量高的材料协调, 但其单一性状突出; R1156的表型与普通小麦相似, 是“小麦型”小黑麦的代表, 其株高仅为87.0 cm, 分蘖较多(19.0个), 穗粒数与主栽品种相近(54.8粒)。
GISH分析发现, 小穗数最多的R1103和高产系R1162均为六倍体小黑麦(AABBRR), 其染色体组成均为2 n = 42 (28W+14R)。通常情况下, 黑麦基因组DNA在黑麦染色体上会产生明显的端带, 如R1103中的14条黑麦染色体在一端或两端具有端带(图2-A1), 而R1162的黑麦染色体中有2条没有端带(图2-B1), 说明这2个材料的黑麦染色体不完全一致。Mc-FISH分析结果表明, R1103和R1162中所含的14条黑麦染色体为1R~7R染色体各1对, 其中两条端带有差异的染色体为4R染色体, 并且2份材料中4R染色体的mc-FISH带型基本一致(图2-A2和B2)。因此, R1103和R1162染色体组成均为2 n = 6 x = 42 (14″W+7″R)。此外, 穗粒数最多的R2209也为六倍体小黑麦(图片未显示), 其染色体组成与R1103相同, 并且14条黑麦染色体都具有端带(一端或两端)。
 | 图2 8份小麦-黑麦种质材料的GISH和mc-FISH鉴定A~F依次为小麦-黑麦种质材料R1103、R1162、R2203、R2221、R1147和R1156。A1~F1是以黑麦基因组DNA为探针(绿色), 中国春基因组DNA为封阻的GISH鉴定结果, 其中B1和D1中的十字星号示无端带的2条黑麦染色体, C1和D1中的箭头示1对黑麦的小染色体。 A2~F2是以重复序列p As1 (红色)和p Sc119.2 (绿色)为探针的mc-FISH鉴定结果, C2和D2中的问号示不能确定归属的染色体。Fig. 2 Identification of GISH and mc-FISH of eight wheat-rye derivativesA-F are wheat-rye derivatives R1103, R1162, R2203, R2221, R1147, and R1156, respectively. A1-F1 are GISH results with rye genomic DNA as a probe (green) and Chinese Spring DNA as a blocker. The cross stars in B1 and D1 show the two rye chromosomes without GISH binding pattern, and the arrows in C1 and D1 show a pair of small rye chromosomes. A2-F2 are mc-FISH results on the same metaphase after GISH analysis by p As1 (red) and p Sc119.2 (green) simultaneously. The question marks in C2 and D2 show the undetermined chromosomes. |
GISH分析确定表现高产的R2203和R2221同为八倍体小黑麦(AABBDDRR), 其染色体组成均为2 n = 56 (42W+14R)。R2203中14条黑麦染色体的一端或两端均具有端带(图2-C1), 而R2221的黑麦染色体中有2条染色体没有端带(图2-D1), 说明这两份材料中的黑麦染色体也不完全一致。我们还发现这2份八倍体小黑麦中含有1对黑麦的小染色体(图2-C1和D1), 而上述六倍体小黑麦材料中没有这样的小染色体(图2-A1和B1)。Mc-FISH结果显示, 2份八倍体材料中的14条黑麦染色体分别为1R、2R、3R、4R、6R和7R染色体各1对, 缺少5R染色体, 但却含有1对黑麦的小染色体(图2-C2和D2); 位于小染色体端部和中部的p Sc119.2带型与5R染色体标准带型不完全一致, 与其他染色体的带型也不具有很好的一致性, 所以暂时不能确定其归属(图2-C2和D2)。两条端带有差异的染色体同样为4R, R2221的4R染色体短臂端部的绿色条带不明显(图2-D2), 可能是由于杂交过程中一些因素所致, 也可能是染色体DNA发生了变异。
GISH鉴定未发现分蘖数最多的R1147 (图2-E1)和千粒重最大的R1017 (图片未显示)中有可见外源黑麦染色体或染色体片段, 利用探针p As1 (红色)和p Sc119.2 (绿色)对R1147的mc-FISH鉴定也未发现其染色体上含有异于标准带型的信号(图2-E2)。
“小麦型”材料R1156的染色体总数为42条, 有1对小麦染色体的短臂被黑麦染色体臂代换, 形成小麦-黑麦易位系(图2-F1)。进一步的mc-FISH分析发现, 该材料中黑麦染色体的1RS代换了小麦染色体的1BS, 所以该易位为T1RS·1BL (图2-F2)。并且, 根据带型可以看出, 所有染色体均成对出现, 因此认为R1156的染色体组成为2 n = 42 (20″W+ 2T1RS·1BL)。
小麦产量构成三要素中分蘖数的贡献最大, 但在高肥力条件下, 分蘖数接近高限, 因此穗粒数和千粒重成为进一步提高小麦产量的限制因素[ 25]。产量构成因素的改进潜力在不同地区可能有所差异, 但通过增加穗粒数和改良株型等在小麦产量提高方面仍具有较大潜力; 要通过与多小穗的近缘种质或其后代杂交、回交, 选育穗略长、小穗略多(如增加1~3个)、穗粒数和粒重协调发展的类型, 即兼顾穗数的大穗大粒高产类型[ 26]。本研究中, 390份小麦-黑麦种质材料的穗长和分蘖数较10个主栽品种有明显优势, 小穗数、穗粒数和千粒重也不同程度地优于主栽品种, 说明黑麦分蘖力强、小穗数多等优良性状已转移到这些远缘杂交材料中。分蘖数、穗长、小穗数、穗粒数、千粒重和株高在390份小麦-黑麦种质材料中均呈连续分布, 并且极差较大, 体现出丰富的多样性。分蘖数、穗长、小穗数、穗粒数和千粒重5个性状均有远高于主栽品种的材料, 有的材料产量相关性状表现比较协调, 籽粒产量较高, 因此, 充分发掘这些小麦-黑麦种质材料的优良性状, 并将其进一步导入到小麦品种中, 对小麦育种工作具有重要意义。近年来, 本实验室已经将农艺性状表现优异的部分材料与生产上的主栽品种杂交, 配制了245个组合, 并进行了回交, 以广泛应用于小麦育种[ 3]。
利用GISH和mc-FISH技术对农艺性状表现突出的小麦-黑麦种质材料进行鉴定, 明确了它们的染色体组成, 这不仅可以为育种家提供遗传背景清楚的育种材料, 也为这些优异材料的进一步研究和利用奠定了基础。本研究中8份代表性材料不仅包括六倍体和八倍体小黑麦, 还包括小麦-黑麦易位系和不携带明显黑麦染色体片段的材料, 且2份八倍体小黑麦(R2203和R2221)与3份六倍体小黑麦(R1162、R1103和R2209)携带不完全相同的黑麦染色体, 八倍体小黑麦中缺少黑麦的5R染色体, 而含有1对黑麦的小染色体(图2-C2和D2), 而六倍体小黑麦中没有这样的小染色体, 其黑麦染色体为1R~7R (图2-A2和B2)。另外这些六倍体和八倍体小黑麦中黑麦4R染色体的GISH端带情况也不相同(图2-A1、B1、C1和D1), 这可能是因为黑麦本身的来源不同, 或是黑麦的品种不同, 也可能是黑麦染色体导入小麦后发生剧烈变化, 染色体结构重排所致[ 27, 28]。
小黑麦是小麦与黑麦经远缘杂交形成的典型双二倍体, 它结合了小麦和黑麦的优良特性, 与小麦杂交成功率更高, 所以常作为黑麦优良基因向小麦转移的桥梁亲本[ 9]。利用八倍体或六倍体小黑麦与普通小麦杂交可以得到异附加系、异代换系和易位系等染色体工程材料。本研究调查的390份小麦-黑麦种质材料的分蘖数、穗长和小穗数等性状具有显著优势, 8份农艺性状优良的材料经分子细胞遗传学鉴定, 有5份为小黑麦, 其中3份为六倍体小黑麦, 2份为八倍体小黑麦。结合农艺性状表型分析来看, 390份小麦-黑麦种质材料大部分可能为六倍体和八倍体小黑麦。八倍体小黑麦与普通小麦杂交在F2和F3代就可形成相当数量的在育种学上具有利用价值的如易位、代换和附加等材料[ 29]。已利用八倍体小黑麦和普通小麦杂交, 在F5代得到1R~7R的单体附加系, 以及1R和2R的二体附加系; 抗病性鉴定发现, 5R和6R单体附加系对白粉病表现免疫[ 30, 31]。利用六倍体小黑麦与普通小麦品种杂交, 已选育出高抗白粉病的1RS·1BL易位系84059和84111以及1R(1B)和1R(1D)代换系[ 32]。利用六倍体小黑麦与普通小麦杂交, 通过表型鉴定选择优异材料, 再利用GISH、FISH技术和分子标记技术跟踪和鉴定黑麦遗传物质的传递, 在F2和F3代获得了一定比例的携带黑麦染色质的材料, 这些材料对大麦黄矮病表现高抗[ 33]。本实验室在小麦-黑麦种质材料与主栽品种杂交的245个组合的后代中也获得一定数量的附加系, 代换系和易位系等材料, 有些材料表现出超亲的优良性状。
为了将这390份小麦-黑麦种质材料应用于小麦育种以丰富小麦的遗传变异, 可以将染色体工程方法与常规育种技术相结合, 在小麦-黑麦种质材料与小麦主栽品种的杂交、回交后代中, 筛选具有目标性状的后代材料, 进一步利用GISH与FISH以及分子标记技术跟踪和检测杂交后代中的黑麦遗传物质, 创制具有优良性状的新种质或新品种。另外, 导入小麦的整条或大片段黑麦染色体, 在引入黑麦优异性状的同时, 往往具有连锁累赘和遗传上不稳定等不利因素, 可利用离子辐射、诱导部分同源染色配对、组织培养和杀配子染色体(gametocidal chromo-some)等方法处理小黑麦或小黑麦与小麦杂交得到的后代(包括附加系、代换系和大片段易位系等), 创制新的小片段的小麦-黑麦易位系, 以有目的地将含有黑麦有益基因的染色体片段导入小麦, 有效利用其优异基因。此外, 由于小黑麦同时具有小麦和黑麦不同属的染色体, 因而是研究异属染色体配对行为及相应性状遗传的理想材料[ 8]。任正隆等[ 29]即利用八倍体小黑麦与普通小麦杂交的后代分析了小麦和黑麦染色体在小黑麦与小麦杂交的不同世代群体中的传递规律。
与六倍体和八倍体小黑麦相比, 易位系携带的非目标遗传物质较少, 所以在所有的小麦-黑麦异染色体系中, 易位系被认为是向小麦中转移外源有益基因的最有效方式。本研究发现R1156为小麦-黑麦的1RS·1BL易位系。Kim等[ 34]通过研究不同来源的黑麦1R染色体的农学遗传效应, 发现不同来源的1RS易位系在籽粒产量上表现显著的遗传多样性, 而小麦遗传背景的效应是影响1RS·1BL易位系农艺性状的主要因素。虽然在利用黑麦优良性状的同时, 一些影响小麦品质的不利性状也会引入小麦[ 35], 但这些不利性状是否导入同样取决于特定的易位系、黑麦染色质的来源和遗传背景等的影响[ 34, 36], 可以通过选择合适的材料尽量将其避免。周建平等[ 37]和任天恒等[ 38]利用绵阳11与不同来源的1RS·1BL易位系杂交, 不仅培育出兼抗条锈和白粉病的小麦品种, 而且新品种表现“延绿”的特性。樊小莉等[ 39]利用小偃6号与德国白黑麦的远缘杂交后代, 选育出高抗条锈病的1RS·1BL易位系。所以可以发掘不同来源的黑麦1RS染色体的遗传多样性, 或利用小麦和黑麦种内的遗传变异, 创造较多的不同种系来源的同类易位体, 从中选育出一个与小麦遗传背景有最佳重组和最佳相互作用的易位材料以应用于小麦育种[ 40]。本研究的结果表明, T1RS·1BL易位系R1156分蘖数多, 综合农艺性状优良, 是一个可以直接应用于小麦育种的易位系。此外, 未检测到 R1147和R1017携带黑麦染色体或染色体片段, 这可能是因为黑麦染色体片段太小, 实验未能分辨出, 或者是完全不携带黑麦遗传物质, 有待利用特异分子标记等技术进一步验证。R1147分蘖数高达32.8, 显著高于主栽品种; R1017千粒重高达60.5 g, 也显著高于主栽品种, 与六倍体、八倍体小黑麦相比, 这两个材料的优良性状更容易转移到普通小麦中。
系统调查了390份小麦-黑麦种质材料的产量相关农艺性状, 发现远缘杂交后代不仅具有丰富的多样性, 而且比主栽小麦品种具有明显的表型优势, 尤其是穗长和分蘖数, 另外小穗数、穗粒数和千粒重方面也具有一定优势。筛选出8份农艺性状优良的小麦-黑麦种质材料, 其在染色体水平上也具有丰富的遗传多样性。本研究结果为这些材料应用于小麦育种提供了依据。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|