
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
一年生簇毛麦α-醇溶蛋白基因的分离、原核表达与功能鉴定
[杨帆1 , 陈其皎1, 2 , 高翔1, 2, *  , 赵万春
, 赵万春1, 2, * , 强琴琴1 , 吴丹1 , 孟敏1 ]
, 赵万春, 强琴琴|
|


醇溶蛋白是面筋的主要成分之一, 对小麦品质具有重要影响。根据数据库中全长α-醇溶蛋白基因设计了1对通用引物, 从5份一年生簇毛麦(
Gliadin, which has a great effect on wheat quality, is one of main components in gluten. According to the full lengths of α-gliadin genes deposited in NCBI database, a conserved primer pair was designed to clone α-gliadin genes in five
小麦是我国主要粮食作物之一, 品质改良是小麦的重要育种目标。但是在普通小麦中发现的能够编码优质蛋白的基因并不多, 并且由于现代农业导致小麦品种越来越单一化, 使得种内遗传变异逐渐丧失, 造成新选育的小麦品种遗传基础越来越狭窄, 限制了小麦品质的进一步改良。因此, 通过在小麦近缘种属中寻找新的优质基因资源来拓宽小麦的遗传基础成为小麦品质育种的一条有效途径。
一年生二倍体簇毛麦[ Dasypyrum villosum, 2 n=14, VV (syn. Haynaldia villosa)], 原产于地中海东北部和高加索地区, 是小麦遗传改良的重要种质资源[ 1, 2, 3]。一年生簇毛麦具有许多重要的小麦病害抗性基因资源, 如抗黑穗病( Ustilago nuda和 U. triticii)[ 4]、白粉病( Erysiphe graminis)[ 5, 6]、纹枯病( Pseudocerc - osporella herpotricoides)[ 7]、锈病( Puccinia graminis和 P . recondite)[ 8]和根腐病( Rhizoctonia solani)[ 9]等多种病害, 同时也具有耐盐[ 10]等抗逆特点。一年生簇毛麦的籽粒蛋白含量高, 在1V、6VS和7V染色体上, 已经发现含有编码种子储藏蛋白的基因位点 Glu-V1[ 11]、 Glu-V3[ 12]、 Gli-V2[ 13]和 Wsp-1[ 14]。此外, 一年生簇毛麦中还携带矮秆[ 15]、产量[ 16, 17]和面筋强度[ 18]的相关基因。
小麦胚乳储藏蛋白是决定小麦品质, 尤其是小麦加工品质的主要因素。它主要由麦谷蛋白(glutenins)和麦醇溶蛋白(gliadins)组成, 为面筋的主要成分, 是决定面团黏弹性的主要因素[ 19]。麦醇溶蛋白约占小麦胚乳贮藏蛋白总量的50%~60%[ 20, 21], 其中α-醇溶蛋白占总醇溶蛋白的25%[ 22]。它包含信号肽(signal peptide)、N-端重复区(N-terminal repetitive region)、多聚谷氨酰胺I区(polyglutamine repeats I)和N-端非重复区(unique non-repetitive domain I), 最后是多聚谷氨酰胺II区(polyglutamine repeats II)和C-端非重复区(unique non-repetitive domain II)[ 23]。目前, 已经从小麦及其近源物种中分离了许多α-醇溶蛋白基因, 但国内外尚未报道关于一年生簇毛麦中α-醇溶蛋白基因对小麦品质贡献的研究。本研究利用简并引物分离一年生簇毛麦中的α-醇溶蛋白基因, 对克隆得到的部分α-醇溶蛋白基因进行原核表达、亲和层析及体外功能鉴定, 为研究一年生簇毛麦中α-醇溶蛋白亚基对小麦品质改良的作用提供基础。
用微量CTAB法[ 24]提取一年生簇毛麦TA10220、TA10224、TA10227、TA10232和TA10249 (由美国堪萨斯州大学小麦遗传与基因组资源中心Bikram S. Gill博士实验室提供)的基因组DNA。利用Primer Premier 5, 根据GenBank (http://www.ncbi. nlm.nih.gov/)中已注册的α-醇溶蛋白基因序列的保守区域设计扩增基因全长编码区的引物(al-GliF: 5′-ATG AAG ACC TTC CTC ATC TTT GTC-3′; al-GliR: 5′-CCA TGT TTG AAC TAG TAT AGG TCG G-3′)。PCR体系为 5 × PrimeSTAR buffer (Mg2+plus) 5 µL, 2.5 mmol L-1dNTP Mixture 2 µL, 0.1 mmol L-1上下游引物各1 µL, 模板DNA 100 ng, 2.5 U µL-1PrimeSTAR HS DNA Polymerase (TaKaRa) 0.2 µL, 加ddH2O至反应体积为25 µL。PCR程序为 94℃ 5 min; 94℃ 45 s、50℃ 50 s、72℃ 90 s, 30个循环; 72℃ 8 min。PCR扩增产物经1.5%琼脂糖凝胶电泳检测后, 切胶回收目的片段。用 pEASY-T1载体连接目的片段并转化大肠杆菌 Trans1-T1 (北京全式金生物技术有限公司), 在选择性培养基上过夜培养, 画线后从每个克隆挑选3个独立阳性菌落送至南京金斯瑞生物科技有限公司测序。
用DNAMAN (6.0)推导分析α -醇溶蛋白的序列; 采用NCBI在线BLAST (http://blast.ncbi.nlm.nih.gov/)进行序列的同源性比对; SignalP 4.1 Server在线完成氨基酸序列信号肽分析; 蛋白质二级结构的预测采用PSIPRED server (http://bioinf.cs.ucl.ac.uk/psipred/); 以MEGA 5.0构建系统演化树; 参照van Herpen等[ 25]和李光蓉等[ 26]报道的方法分析识别α -醇溶蛋白序列的毒性多肽位点。
设计1对表达引物Ex.al-gliF (5′-GTA CTT AGA GTT CCA GTG CCG C-3′)/Ex.al-gliR (5′-TTA GTT ACT ACC GGT GCT ACC AAA T-3′), 分别对含有2条代表性序列的阳性质粒进行PCR扩增(KJ004708和KJ004714), 回收目的片段并分别连接至表达载体 pEASY-E1, 转化表达菌株BL21 (DE3)(北京全式金生物技术有限公司), 挑取阳性菌落接种至LB中(氨苄青霉素, 100 mg mL-1), 37℃过夜培养后按照体
积比1∶100将培养物接种到新鲜LB中培养至 A600 = 0.5~0.8, 取1 mL过夜培养的菌液作为阴性对照, 向剩余的菌液中加入诱导剂IPTG (终浓度1 mmol L-1)诱导6 h (37℃, 200转 min-1)。9300 × g离心10 min收集菌体, 加入1 × SDS凝胶加样缓冲液重悬, 煮沸7 min, 以12% SDS-PAGE电泳检测蛋白的表达, 并用消毒手术刀在PAGE胶上切下与预测分子量相符的正确表达的蛋白, 采用MALDI-TOF/TOF 4800 (Applied Biosystem)质谱仪进行串联质谱鉴定(中国科学院上海生命科学研究院蛋白质组研究分析中心), 采用一级肽指纹质量与二级肽碎片质量综合分析法Combined (MS+MS/MS)分析样品序列, 通过NCBI (http://www.ncbi.nlm.nih.gov/)序列同源性比对确定蛋白质的性质。
离心收集诱导后的菌体(4℃, 6800 × g, 10 min), 加入破碎缓冲液(50 mmol L-1 Tris-HCl, 100 mmol L-1NaCl, 2 mmol L-1 EDTA, 1 mg L-1溶菌酶, pH 8.5)在冰上放置45 min, 超声破碎细胞并离心收集沉淀(4℃, 15 000 × g, 10 min)。沉淀经6 mol L-1盐酸胍的磷酸盐缓冲液预处理后, 采用 ProteinIso Ni-NTA Resin (北京全式金生物技术有限公司)纯化和收集目的蛋白。将收集的蛋白透析72 h, 低温冷冻干燥48 h后在-20℃冰箱中保存备用。
采用4 g微量粉质仪测定一年生簇毛麦α-醇溶蛋白亚基的品质效应。在4 g基础面粉中加入50 mg纯化蛋白和适量水, 混合4 min后加入0.25 mL DTT溶液(50 mg L-1)反应1 min, 再加入0.25 mL KIO3溶液(200 mg L-1)反应10 min, 记录粉质曲线和相应品质参数。
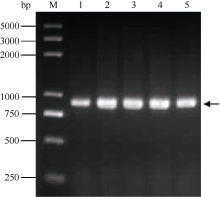
通过分析已公布的α -醇溶蛋白基因, 利用序列的保守区设计1对简并引物, 以一年生簇毛麦基因组DNA为模板扩增得到52条序列(GenBank登录号为KJ004676~KJ004727)。NCBI在线BLAST表明, 得到的序列与已公布的多年生簇毛麦α -醇溶蛋白基因序列相似度较高, 最高可达94%。克隆得到的序列全长介于816~873 bp之间(图1), 最常见的基因片段长度为858 bp, 共出现19次。
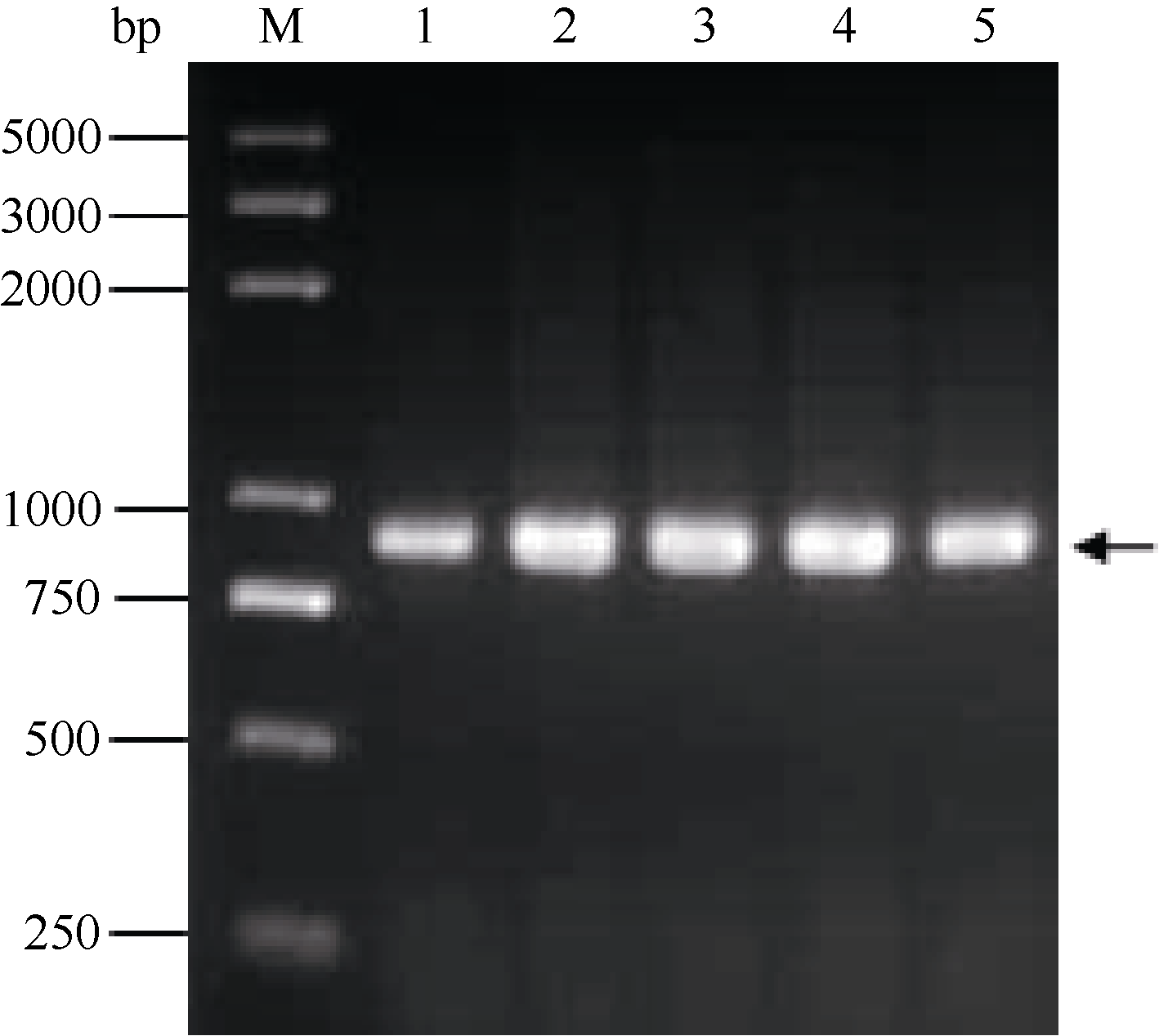 | 图1 1.5%琼脂糖凝胶扩增α-醇溶蛋白基因序列Fig. 1 Amplification products of α-gliadin gene sequences separated on 1.5% agarose gelM: Trans 2K Plus DNA marker; 1: TA10220; 2: TA10224; 3: TA10227; 4: TA10232; 5: TA10249. |
DNAMAN开放阅读框分析表明, 8条序列(KJ004720~KJ004727)含有提前终止密码子, 属于假基因。其中KJ004725在642 bp处(特征区II)缺失碱基C导致阅读框移码, 其他假基因均因碱基替换所致。碱基替换在N端重复区出现的频率最高(3/7)。在7个碱基替换的类型中, 有6个均因密码子的第1位碱基C被T替换, 分别导致CAA→TAA (3/6)和CAG→TAG (3/6), 而KJ004726是因密码子的后两位碱基同时被碱基A替换导致TGG→TAA。KJ004680则因3′端倒数第8个密码子的第3位缺失碱基T而移码突变, 使得终止密码子缺失。
SNP分析表明, 碱基转换占73%, A/G和T/C类型分别占39%和34%; 碱基颠换只占27%, 并以A/T颠换为主(58%), 其他几种碱基颠换的比例较低。
分析推导的氨基酸序列, 得到43条完整的α-醇溶蛋白序列, 编码271~290氨基酸残基, 均具有典型的α-醇溶蛋白结构, 分别是由19个氨基酸残基组成的信号肽, N-端重复区, 多聚谷氨酰胺I区, 特征区I, 多聚谷氨酰胺II区和特征区II。根据N-端重复区多肽序列的差异, 将这43条α-醇溶蛋白分为5种类型(图2-A)。序列比对分析发现, 氨基酸变异位点大部分存在于N-端、特征区I和特征区II, 导致Gln和Arg、Leu和Pro, 以及Gln和Leu之间的相互转换。N-端重复区和多聚谷氨酰胺Ⅰ区多肽长度的变异主要是由于第V类亚基中的4个α-醇溶蛋白(KJ004681、KJ004693、KJ004709和KJ004711)缺失了富含Gln的片段。本试验获得的一年生簇毛麦α-醇溶蛋白多聚谷氨酰胺I区的长度介于11~23个氨基酸残基之间, 其中Gln的平均含量为18.1个。有33条序列发生了Q→E突变, 其中KJ004682还发生了Q→R突变。多聚谷氨酰胺II区序列都非常短, 只含有5个氨基酸残基(QQLQQ)且非常保守(图2-B), 只有KJ004717在这个区域的第1位氨基酸残基由谷氨酰胺Q变异为亮氨酸(LQLQQ)。
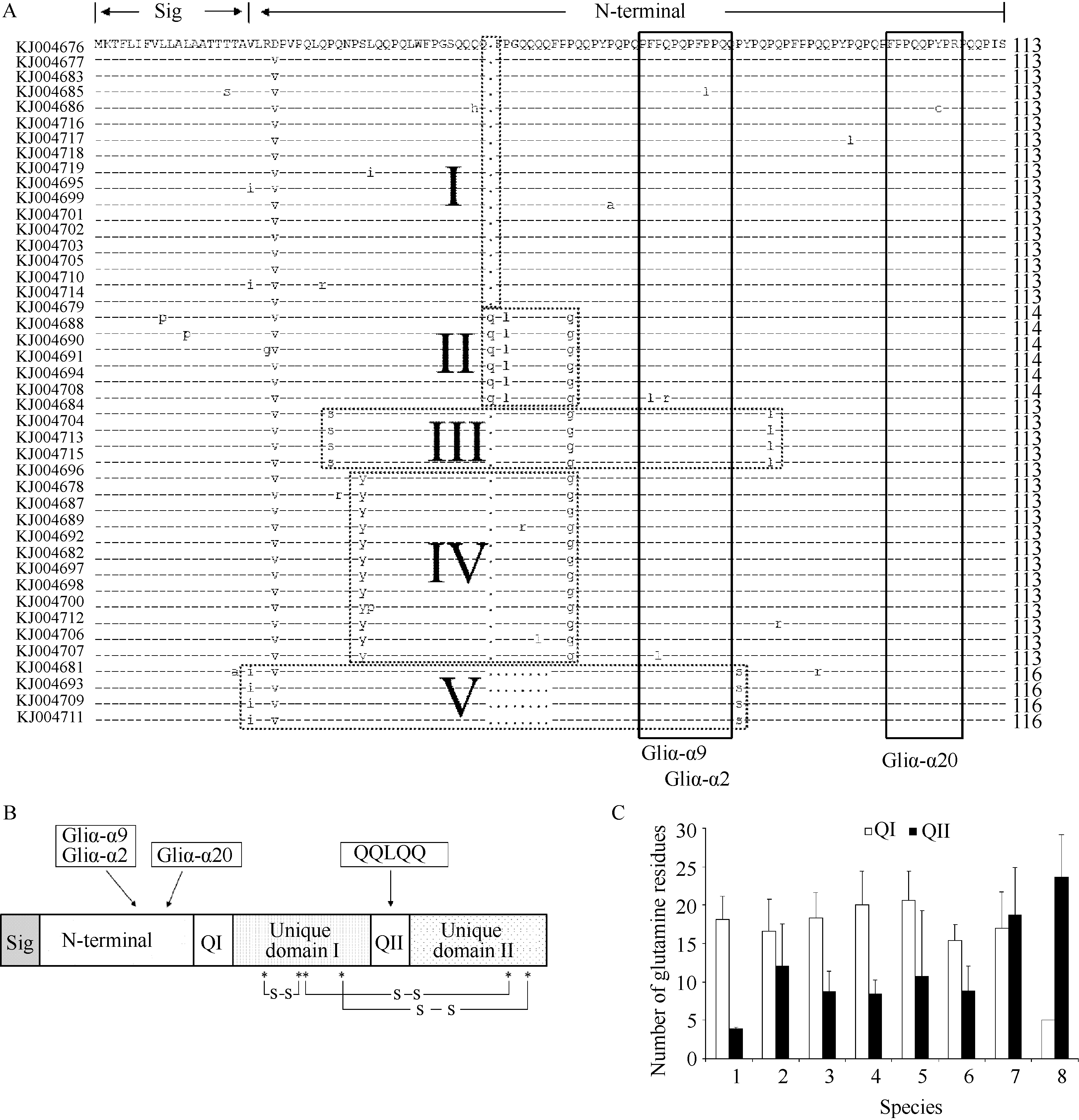 | 图2 α-醇溶蛋白基因推导氨基酸序列分析A: 43条推导氨基酸序列的毒性多肽位点和分类, 粗线框示毒性多肽位点, 虚线框示不同类型的α-醇溶蛋白。B: α-醇溶蛋白的结构及二硫键模式, N端重复区(N-terminal)存在毒性多肽位点, 多聚谷氨酰胺II区(Q II)存在QQLQQ多肽序列, 星号(*)示半胱氨酸残基的位置。C: 不同物种α-醇溶蛋白在多聚谷氨酰胺I区和II区中谷氨酰胺残基数量的变异, 误差线表示标准差。1: 一年生簇毛麦, n= 43; 2: 普通小麦, n=26; 3: 粗山羊草, n= 16; 4: 一粒小麦, n= 15; 5: 二粒小麦, n= 17 ; 6: 黑麦, n= 17; 7: 拟斯卑尔脱山羊草, n= 7; 8: 稃旱麦草, n= 7。Fig. 2 Deduced amino acid sequence analysis of α-gliadin genesA: T-cell stimulatory epitopes and classification of 43 α-gliadin amino acid sequences, thick frames indicate T-cell stimulatory epitopes, dot line frames indicate different types of α-gliadins. B: Schematic structure of α-gliadin and intra-molecular disulfide bonds, T-cell stimulatory epitopes existed in the N-terminal, QQLQQ existed in the Q II, * indicates the positions of cysteine residue. C: Variations in mean number of glutamine residues in the polyglutamine domains (Q I and Q II) of α-gliadins among different species. Error bars show the standard deviations, 1: Dasypyrum villosum, n= 43; 2: Triticum aestivum, n= 26; 3: Aegilops tauschii, n= 16; 4: Triticum monococcum, n= 15; 5: Triticum d i coccoides, n= 17; 6: Secale cereale, n= 17; 7: Aegilops speltoides, n= 7; 8: Er e mopyrum bonaepartis, n= 7. |
通过比较不同物种间多聚谷氨酰胺I和II区中谷氨酰胺残基数量的变异, 发现不同物种间多聚谷氨酰胺区中Gln的数量变异比较大。除拟斯卑尔脱山羊草和稃旱麦草外, 其他6个物种多聚谷氨酰胺I区中Gln的数量要多于多聚谷氨酰胺II区。与其他物种相比, 本研究从一年生簇毛麦中获得的α-醇溶蛋白在多聚谷氨酰胺II区中Gln数量的变异较小(图2-C)。
α-醇溶蛋白中Cys的位置和数目都非常保守, 分别在特征区I和特征区II中含有4个和2个Cys (图2-B)。在所得到的序列中, KJ004677、KJ004686、KJ004696和KJ004714含有7个Cys。除KJ004696的额外Cys是由Ser变异而来, 其他3个序列的额外Cys均由Tyr变异而来(Tyr→Cys)。进一步分析额外Cys出现的位置, 发现其出现在KJ004686的N-端重复区、KJ004696的特征区I、KJ004677和KJ004714的特征区II。
α-醇溶蛋白中主要含有4个能诱发乳糜泻病的多肽因子, 分别是Gliα-α (QGSFQPSQQ)、Gliα-α2 (PQPQLPYPQ)、Gliα-α9 (PFPQPQLPY)和Gliα-α20 (FRPPQQPYPQ)[ 25, 26], 人体T细胞与这些特定多肽结合后会引起小肠黏膜损伤和吸收不良[ 27]。不同的诱发因子有特定的保守分布区域, Gliα-α出现在特征区II, 而Gliα-α2、Gliα-α9和Gliα-α20均分布在N-端重复区, 且Gliα-α2和Gliα-α9多肽序列有部分重叠(图2-A)。将43条一年生簇毛麦α-醇溶蛋白多肽序列与这4个毒性多肽位点对比分析表明, 所有序列中都不含有完整的Gliα-α2、Gliα-α9和Gliα-α20毒性多肽位点序列, 并缺失Gliα-α位点(表1)。
| 表1 不同研究获得的一年生簇毛麦α-醇溶蛋白毒性多肽位的比较 Table 1 Comparison of T cell stimulatory epitopes of α-gliadins in Dasypyrum villosum obtained from different studies |
比较发现, 不同物种的α-醇溶蛋白毒性多肽位点分布差异非常大(表2)。其中冰草属和本研究中的5份一年生簇毛麦品系均不含4个毒性多肽位点, 旱麦草属只含有Gliα-α位点, 黑麦属缺失Gliα-α20位点, 而偃麦草属、山羊草属和小麦属则含有全部4个毒性多肽位点(表2)。
| 表2 不同物种α-醇溶蛋白中毒性多肽位点数量的比较 Table 2 Comparison of the quantity of T cell stimulatory epitopes in α-gliadins among different species |
一年生簇毛麦α-醇溶蛋白的二级结构主要由α-helix和coil组成。α-helix出现的位置和数目比较保守, 在特征区II中除KJ004696含有2个α-helixes外, 其他均只含1个α-helix, 且在N-端重复区不含α-helix。成熟的一年生簇毛麦α-醇溶蛋白中α-helix通常出现5次, 但是KJ004677、KJ004679、KJ004682、KJ004685、KJ004694、KJ004696、KJ004700、KJ004704、KJ004713、KJ004715和KJ004716中有6个α-helixes, 另外KJ004683和KJ004702中只含4个α-helixes。在这43条α-醇溶蛋白中, 除KJ004685、KJ004691、KJ004697、KJ004707和KJ004713在特征区II出现一次β-strand (分别占序列的1.1%、0.8%、1.1%、0.7%和0.8%)外, 其他序列均不含β-strand。
| 表3 原核表达的α-醇溶蛋白串联质谱鉴定 Table 3 Identification of prokaryotically expressed α-gliadins using MALDI-TOF/TOF tandem mass spectrometer |
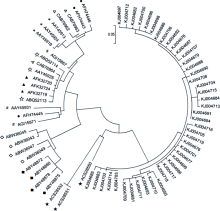
将获得的43条一年生簇毛麦α-醇溶蛋白序列与NCBI中已注册的小麦以及其他近缘种的α-醇溶蛋白氨基酸序列构建系统进化树, 发现本实验中得到的所有一年生簇毛麦α-醇溶蛋白被单独聚为一类, 与小麦及小麦其他近源物种的α-醇溶蛋白亲缘关系较远, 同时与Li等[ 28]得到的一年生簇毛麦亲缘关系也较远, 推测本研究从这5份一年生簇毛麦品系中获得的α-醇溶蛋白为醇溶蛋白家族新成员(图3)。
 | 图3 α-醇溶蛋白序列的系统演化树分析KJ004676~KJ004719 (不含KJ004680)来自本研究。■: 一年生簇毛麦; ◇: 多年生簇毛麦; #: 拟斯卑尔脱山羊草; ☆: 粗山羊草; ★: 华山新麦草; ▲: 黑麦; ▽: 普通小麦; *: 一粒小麦; : 二粒小麦。Fig. 3 Phylogenetic analysis of the α-gliadinsKJ004676 to KJ004719 (except KJ004680) were from this study. ■: Dasypyrum villosum; ◇: Dasyp y rum breviaristatum; #: Aegilops speltoides; ☆: Aegilops tauschii; ★: Psathyrostachys huashanica; ▲: Seca le cereal; ▽: Triticum aestivum; *: Triticum monococcum; : Trit i cum dicoccoides. |
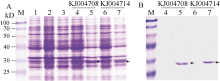
选取两条具有代表性的一年生簇毛麦α-醇溶蛋白序列, 对其进行体外功能鉴定, 其中KJ004708含有α-醇溶蛋白典型的6个半胱氨酸残基, KJ004714在特征区II含有一个额外的半胱氨酸残基。含有重组表达质粒的菌株在37℃经过1 mmol L-1 IPTG诱导6 h后, SDS-PAGE电泳检测结果表明在约30 kD处出现目的条带, 与预测的分子量大小相符(图4-A)。将表达条带进行切胶和串联质谱测序后(表3), 对质谱鉴定的α-醇溶蛋白肽段序列进行比对表明原核表达产物的氨基酸序列与推导的氨基酸序列完全一致, 且与NCBI数据库来源的α-醇溶蛋白相似度高于60%, 但不同来源的α-醇溶蛋白在多肽片段DVLLQQHNI/VAPIR中的序列差异很大(表3)。表达产物纯化后经SDS-PAGE电泳分析表明在约30 kD处同样出现明显目的条带, 证明原核表达产物纯化成功(图4-B)。
 | 图4 蛋白表达和纯化的SDS-PAGE分析A: 诱导表达产物的SDS-PAGE分析; B: 纯化产物的SDS-PAGE分析。M: Blue Plus protein marker; 1: 未诱导的表达菌株(DE3); 2: 未诱导空载体; 3: 诱导空载体; 4和6: 未诱导的重组质粒; 5和7: 诱导后的重组质粒。箭头示目的蛋白。Fig. 4 SDS-PAGE analysis of protein expression and protein purificationA: analysis of expressed proteins by SDS-PAGE; B: analysis of purified proteins by SDS-PAGE. M: Blue Plus protein marker; 1: protein of BL21 (DE3) expression strain without induction; 2, protein of non-recombinant plasmid without induction; 3, protein of non-recombinant plasmid after induction; 4 and 6, protein of recombinant plasmid without induction; 5 and 7, protein of recombinant plasmid after induction. Target proteins are indicated by arrows. |
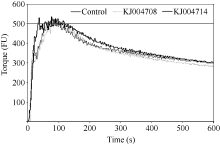
采用4 g微量粉质仪通过氧化还原反应分别将50 mg纯化后的表达产物掺入到基础面粉中, 通过测定面团的粉质曲线, 分析粉质参数的变化来揭示一年生簇毛麦中不同类型的α-醇溶蛋白亚基对小麦品质的影响。分析表明, 添加KJ004708亚基后面团的形成时间、弱化度、公差指数和粉质质量指数都有显著增加, 而添加KJ004714亚基后面团的7个主要的粉质参数较对照均出现显著变化(图5和表4)。
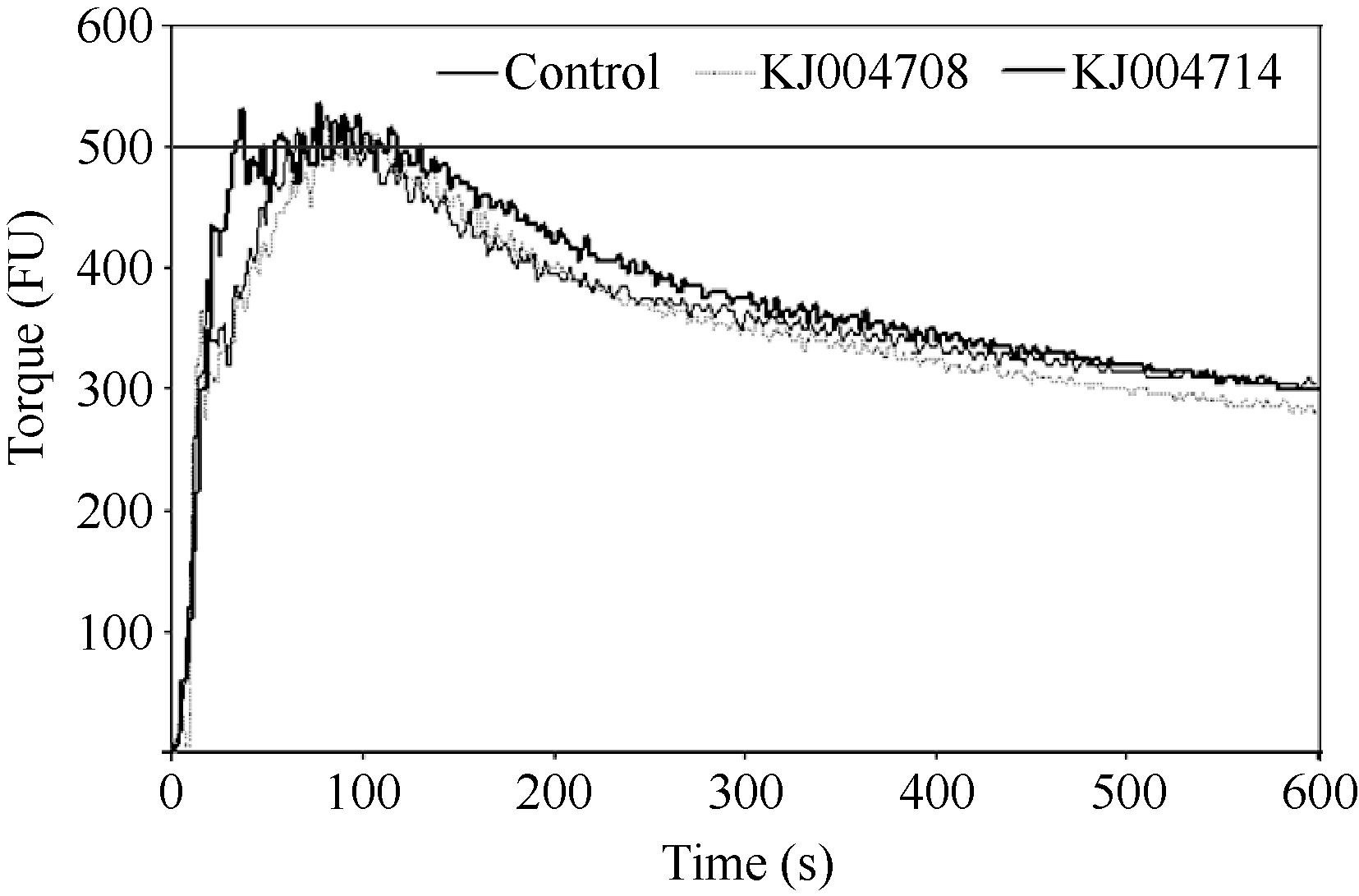 | 图5 纯化的α-醇溶蛋白亚基对粉质曲线的影响Fig. 5 Effects of purified α-gliadin subunits on the Farinograph parameters |
| 表4 添加KJ004708和KJ004714对小麦面团粉质特性的影响 Table 4 Effects of KJ004708 and KJ004714 on Farinograph parameters |
一年生簇毛麦中不仅含有许多重要的抗逆位点, 而且还具有能增加籽粒蛋白含量和增强面筋强度等优质性状, 是小麦品种改良的重要遗传资源。目前已经培育了许多不同的小麦-簇毛麦代换系、易位系等材料, 有效地拓宽了小麦的遗传资源并加快了小麦育种进度。目前已经从不同的物种中分离得到了许多α-醇溶蛋白基因, 但是假基因比例都比较高, 如van Herpen等[ 25]从小麦不同的染色体祖先中获得的230条α-醇溶蛋白基因中假基因的比率就高达87%, 其他的一些研究者获得的α-醇溶蛋白基因也存在比较高的假基因频率[ 30, 31, 32]。据推测, α-醇溶蛋白基因家族中假基因出现的频率高达50%[ 33], 而本研究从一年生簇毛麦中共获得了52条序列, 其中仅有8条假基因(15.4%), 不仅远低于前人从不同物种中得到的假基因频率, 也低于Li等[ 28]从簇毛麦中得到的假基因比例(31.3%), 推测获得的假基因频率与不同来源的材料有密切关系。醇溶蛋白高比例假基因的发生可能是染色体非均等交换和基因大量复制过程中的突变所致[ 21, 34]。突变产生的类型中, 碱基C被T替换是最常见的类型之一, 一般认为, 由于醇溶蛋白中含有大量Gln, 而编码Gln的密码子CAA和CAG中的第1位C一旦被T替换就会产生翻译的终止, 产生假基因[ 33], 本试验中的6个假基因(75%)属于此类。van Herpen等[ 25]曾报道此类假基因产生比例为77.2%, 而张晓霞等[ 31]研究发现假基因中有95%是C-T突变类型。本研究中C-T突变频率占总突变频率的34%, 仅次于A-G突变(39%), 而最终假基因频率仅有15.4%, 并且主要由于C-T突变造成, 说明这5份一年生簇毛麦品系中较低的假基因频率并不是由于C-T突变频率较低引起的。本研究中还发现一个密码子的后两位碱基被同时替换(TGG→ TAA)而产生终止密码子的例子。
α-醇溶蛋白序列结构具很高保守性, 典型特征是含5个不同的区域(图2-B)。根据N端重复区多肽序列的不同, 将本试验获得的α-醇溶蛋白分为5种类型(图2-A)。研究一般认为多聚谷氨酰胺II区是α-醇溶蛋白氨基酸序列变异比较大的一个区域[ 26, 31, 35], 而本研究得到的序列在此区域不仅非常保守而且仅含4个Gln (图2-B), Li等[ 28]从一年生和多年生簇毛麦中获得的α-醇溶蛋白在此区域分别平均含14.3个和18.6个Gln, 而李光蓉等[ 26]从多年生簇毛麦中得到的α-醇溶蛋白在多聚谷氨酰胺II区也平均含9.1个Gln。贮藏蛋白亚基中Cys的含量和位置会对亚基的品质效应产生较大影响。α-醇溶蛋白中Cys的数目和相对位置非常保守, 一般含6个Cys。其中有4个位于特征区I, 另外2个位于特征区II, 并且第1和第2位, 第3和第5位, 第4和第6位的Cys可以形成3对分子内二硫键[ 36]。本研究共得到4个含额外Cys的序列, 额外的Cys可能参与分子间二硫键的形成, 对稳定蛋白空间网络结构和改善面筋品质具有积极作用。在这4条序列中只有KJ004696是由于Ser突变为Cys, 其余3条均由于Tyr突变为Cys, 其中KJ004686在N-端重复区倒数第9位密码子后2位碱基同时发生转换突变(TAC→TGT), 导致Tyr→ Cys突变。其他的一些研究也发现在α-醇溶蛋白中存在一个额外的Cys, 但这些额外的Cys多出现在特征区II并且是由丝氨酸(Ser)变异产生的[ 22, 34, 37, 38]。Li等[ 28]从一年生簇毛麦中分离了一个在特征区I含一个额外Cys的α-醇溶蛋白, 但这个Cys仍由Ser变异而来。
聚类分析发现, 本研究得到的一年生簇毛麦α-醇溶蛋白被单独聚为一类, 同其他物种的亲缘关系较远。Li等[ 28]通过分离一年生和多年生簇毛麦中的α-醇溶蛋白并构建系统进化树, 发现簇毛麦属和小麦属间的亲缘关系较远, 同时一年生和多年生簇毛麦α-醇溶蛋白也分别被单独聚为一类, 与本研究得出的结果相似, 推测簇毛麦中α-醇溶蛋白是一个新的α-醇溶蛋白家族。同时本研究中获得的α-醇溶蛋白与其他一年生簇毛麦来源的α-醇溶蛋白[ 28]亲缘关系也较远, 说明不同来源的一年生簇毛麦中α-醇溶蛋白变异非常丰富。α-醇溶蛋白中对乳糜泻病人具有毒性的多肽主要有4种, 分别是Glia-α、Glia-α2、Glia-α9和Glia-α20。序列比较结果表明, 本研究获得的一年生簇毛麦α-醇溶蛋白均不含Glia-α毒性多肽位点, 且由于氨基酸残基的变异导致不含有完整的另外3个毒性多肽位点。Li等[ 28]从一年生簇毛麦中得到的α-醇溶蛋白含有完整的Glia-α位点, 但是缺失Glia-α20毒性位点序列, 另外也不含有完整的Glia-α2和Glia-α9位点(表1)。说明不同来源的一年生簇毛麦中α-醇溶蛋白毒性多肽位点序列变异很大, 与聚类分析的结果相符, 这有助于选育不含有毒性多肽位点的新品种。
微量掺粉试验结果表明, 一年生簇毛麦中两种不同类型的α-醇溶蛋白亚基对小麦面团品质都能产生显著影响。面团的稳定时间、形成时间和断裂时间越长, 带宽值愈大, 同时弱化度和公差指数值越小, 说明面团的面筋强度和弹性越好, 而粉质质量指数则是用于综合评价面粉筋力和耐揉性一个重要依据[ 34, 39, 40]。粉质结果显示, 虽然KJ004708显著增加了弱化度和公差指数, 降低了面团的面筋强度, 但是对面团的形成时间和最终的粉质质量指数有显著的提升作用(表4)。含有一个额外Cys的KJ004714亚基显著作用于所有的7个主要粉质参数, 添加KJ004714能够显著增加面团的稳定时间、形成时间、断裂时间、带宽和粉质质量指数, 同时又显著降低面团的弱化度和公差指数, 对面团面筋的强度和弹性都有显著改善(表4)。推测KJ004714中含有的额外Cys参与分子间二硫键的形成, 促进蛋白质亚基间的聚合, 从而显著地改善面团的结构, 推测KJ004714对小麦面粉品质的改良具有很大潜力。李敏等[ 34]对来源于普通小麦的一个含7个Cys的α-醇溶蛋白品质鉴定表明, 加入外源蛋白亚基后面团的断裂时间、稳定时间和粉质质量指数值显著降低, 而弱化度和公差指数值却显著升高, 说明此亚基对面团品质有不利影响。并且本研究和李敏等[ 34]得到的α-醇溶蛋白亚基中额外Cys均出现在特征区II, 但最终对于品质的影响相差非常大, 说明额外Cys对小麦品质的影响可能非常复杂, 不仅与Cys的数目和位置有关, α-醇溶蛋白本身的序列也可能影响亚基的品质效应。
利用特异引物, 从5份一年生簇毛麦材料中分离得到52条序列, 其中8条为假基因, 1条由于碱基的缺失导致移码突变而缺失终止密码子。得到的43条完整α-醇溶蛋白序列可被划分为5种不同的类型, 序列间的差异主要存在于N端重复区和多聚谷氨酰胺I区。毒性多肽位点的比较、聚类分析和质谱鉴定结果的序列比对分析发现不同来源的簇毛麦α-醇溶蛋白序列间差异较大, 这可能对小麦品质育种产生有利的影响。本研究中获得的α-醇溶蛋白既缺失Gliα-α毒性多肽位点, 也不含完整的其他3个毒性多肽位点。KJ004708与KJ004714均能改善小麦面团的品质, 说明这些一年生簇毛麦品系是小麦品质育种的优良材料, 并且KJ004714对品质的改善更为显著, 其所含额外Cys有助于小麦面筋网络结构的形成。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|