


{kind=link}
{kind=link}
{kind=link}
{kind=link}
灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响
[张英华 , 杨佑明, 曹莲, 郝杨凡, 黄菁, 李金鹏, 姚得秀, 王志敏
, 杨佑明, 曹莲, 郝杨凡, 黄菁, 李金鹏, 姚得秀, 王志敏* ]
, 杨佑明, 曹莲, 郝杨凡, 黄菁, 李金鹏, 姚得秀, 王志敏]
|
|


第一作者联系方式: E-mail:zhangyh1216@126.com
为揭示小麦叶与非叶器官抗氧化系统对灌浆期高温胁迫的反应特征, 探讨不同品种和不同器官耐热性差异机制, 以小麦强耐热品种石家庄8号和弱耐热性品种河农341为材料, 于灌浆期用塑料膜搭棚进行增温处理(花后第8天至第22天), 研究高温胁迫对旗叶光合速率( Pn)、叶绿素含量、旗叶和非叶器官中丙二醛(MDA)和脯氨酸(Pro)含量及超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性的影响。高温处理下, 两品种 Pn比正常温度下(对照)低18.7%~24.9%, 叶绿素含量低5.7%~6.2%; 旗叶、旗叶鞘、穗下节、颖片和籽粒的MDA含量和Pro含量均升高, 其中MDA升高幅度为旗叶>非叶器官, Pro升高幅度为非叶器官>旗叶。旗叶、颖片、籽粒的SOD活性和旗叶、旗叶鞘、籽粒的CAT活性以及旗叶、旗叶鞘、颖片的POD活性在高温胁迫初期即诱导增强, 而其他器官的抗氧化酶活性则在高温持续一段时间后诱导增强, 之后随着高温的持续各器官抗氧化酶活性多表现为低于对照, 高温解除后旗叶鞘、穗下节、颖片的SOD活性和旗叶、颖片、籽粒的POD活性有恢复迹象, 高温对其他器官的SOD和POD活性以及所有器官的CAT活性造成不可逆影响; 总体来看, 非叶器官持续抗氧化能力和耐热性强于叶片。石家庄8号叶与非叶器官细胞膜稳定性、抗氧化酶活性均高于河农341, 显示其整株耐热性强于河农341, 这是石家庄8号在高温胁迫下产量下降幅度低于河农341的重要生理基础。因此认为, 非叶器官在小麦适应灌浆期高温逆境中发挥重要作用。
, YANG You-Ming, CAO Lian, HAO Yang-Fan, HUANG Jing, LI Jin-Peng, YAO De-Xiu, WANG Zhi-Min
Winter wheat in North China has been subject to high temperature stress during grain filling. The purpose of this study was to determine the effect of high temperature on the photosynthetic capability of flag leaf and the antioxidant system of flag leaf and non-leaf organs in winter wheat cultivars Shijiazhuang 8 (tolerant to high temperature) and Henong 341 (sensitive to high temperature). High temperature (HT) stress was imposed with a plastic shed from the 8th to the 22nd day after anthesis, and normal temperature was used as the control. Under HT, the photosynthetic rate and chlorophyll content of flag leaf decreased by 18.7%-24.9% and 5.7%-6.2%, respectively; whereas the malodialdehyde (MDA) and proline (Pro) contents in flag leaf blade, sheath, peduncle, glume and grain increased in different levels. The increased percentage of MDA was higher in flag leaf than in non-leaf organs and that of Pro was higher in non-leaf organs than in flag leaf. Superoxide dismutase (SOD) in flag leaf, glume and grain, catalase (CAT) in flag leaf, sheath and grain, and peroxidases (POD) in flag leaf, sheath and glume were induced at early stage (4 d after treatment) by HT treatment, while the activities of antioxidant enzymes in other organs increased at later stage (7 d after treatment). Thereafter, the activities of antioxidant enzymes in various organs maintained lower levels compared with those of the control. At the 26th day after anthesis when HT stress was relieved for four days, the SOD activity in sheath, peduncle and glume and the POD activity in flag leaf, glume and grain began to increase, but the effects of HT on the SOD and POD activities in other organs and the CAT activity in all organs seemed irreversible. In general, the non-leaf organs exhibited higher antioxidant capability and heat tolerance than flag leaf. Compared with Henong 341, Shijiazhuang 8 exhibited higher cell membrane stability and antioxidant activity in leaf and non-leaf organs, leading to a tolerance to HT stress of the whole plant. This might be the physiological basis of smaller percentage of yield loss in Shijiazhuang 8 than in Henong 341 under HT stress. Our results indicate that non-leaf organs of wheat play an important role in adaptability to climate warming.
我国华北地区小麦生长后期常遇干热风危害[1]。随着全球气候变暖, 高温胁迫对小麦籽粒产量的影响将会更为严重, 因而受到广泛关注[2]。研究表明, 高温胁迫下, 叶片的细胞结构与功能受到破坏, 叶绿素含量明显下降, 光合能力下降, 并最终导致籽粒干物质积累速率降低, 粒重降低[2, 3, 4, 5, 6, 7]。高温对叶片机能的影响程度与细胞内抗氧化系统活性有密切关系[2, 8, 9, 10]。高温诱导叶片中产生过量活性氧, 使膜质过氧化, 加速细胞解体。植物体内具有抗氧化系统, 如抗氧化酶可有效清除植物组织中的活性氧, 对细胞有保护作用。一些学者研究了小麦生育后期高温与膜酯过氧化及抗氧化活性的关系[2, 11, 12], 但现有的研究主要集中在叶片上, 对叶片以外的非叶器官研究较少。据报道, 小麦非叶器官(颖片、叶鞘、节间)光合耐逆性高于叶片[13, 14]。在灌浆期热胁迫处理后, 旗叶叶片光合速率显著下降, 而穗光合下降幅度明显较小, 穗下节间和叶鞘的光合活性也具有相对的稳定性[14]。干旱胁迫后, 穗和芒等非叶器官的光合速率也高于旗叶, 尤其芒在干旱下能提高水分传导, 降低穗温, 对产量有重要贡献[15]。非叶器官对高温、干旱的相对不敏感与其结构、水势稳定性、磷酸烯醇式丙酮酸羧化酶(PEPC)等C4酶活性有密切关系[14, 15, 16, 17]。然而, 叶片与非叶器官间耐逆性的差异是否与抗氧化系统活性有关尚不完全清楚。本文研究了灌浆期高温处理对旗叶光合性能、叶与非叶器官细胞膜稳定性、抗氧化酶活性和有关渗透调节物质含量的影响, 进一步探讨小麦整株不同器官对高温胁迫的反应与适应性差异及其机制, 为小麦抗逆栽培调控和抗逆育种选择提供部分理论依据。
2010— 2011年度在中国农业大学实验站(河北吴桥)进行大田试验, 利用晚播、开花期推迟造成灌浆期高温环境。正常播期(对照)为10月10日, 晚播日期为10月28日。对8个冬小麦品种进行灌浆期耐热性初筛, 从中选出2个耐热性差异大的品种, 即耐热性强的品种石家庄8号耐热性弱的品种河农341, 这2个品种花期基本一致。
2011— 2012年度在同一地点进行石家庄8号和河农341的比较试验, 设正常温度(对照)和大棚增温处理。试验地0~20 cm土层含有机质1.1%、全氮0.12%、速效磷20.9 mg kg-1、速效钾97.1 mg kg-1。10月12日足墒播种, 底施有机肥(鸡粪+土杂粪) 15 m3 hm-2、纯氮157.5 kg hm-2、P2O5 138 kg hm-2、K2O 112.5 kg hm-2。拔节期和开花期各浇水一次, 每次灌水定额750 m3hm-2, 其他田间管理措施同大田生产。每个品种6个小区, 小区面积10 m2。
开花期在各小区选取生长一致的植株作好标记, 开花后第8天, 在各品种3个小区搭建塑料大棚进行增温处理[14], 以不增温小区为对照。大棚面积与小区面积相同, 高度为1.6 m。每天记录棚内外温度, 统计每天9:00、13:30和16:30的温度, 平均值见图1。中午异常高温时揭膜降温, 下午封膜。从花后第8天至花后第22天进行高温处理, 持续15 d, 处理结束后拆棚恢复到与对照一致的温度水平直至成熟。处理与对照区田间湿度和光照差异不显著。
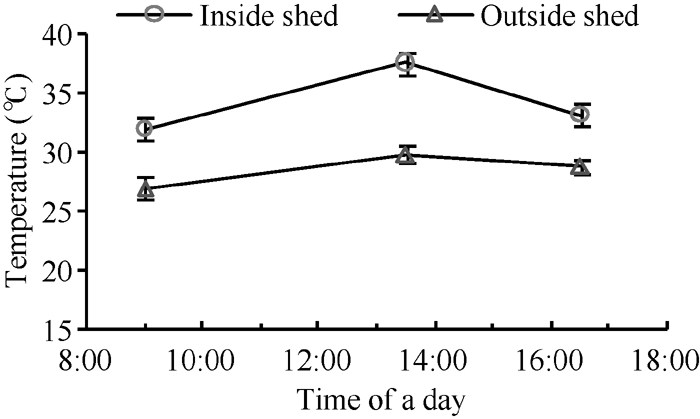 | 图1 大棚内外温度日变化Fig. 1 Diurnal variation of temperature inside and outside the shed |
2011— 2012年度, 于花后8、12、15、18、23、26 d (即处理后0、4、7、10、15 d及恢复后3 d)测定标记植株的净光合速率(Pn)和叶绿素相对含量, 并取旗叶片、旗叶鞘、穗下节、颖片和籽粒, 用液氮速冻后于-80℃冰箱保存, 用于测定丙二醛(MDA)和脯氨酸(Pro)含量及3种抗氧化酶活性。
1.2.1 旗叶Pn和叶绿素相对含量 在每个观测日, 选晴天上午9:30至12:30, 使用LI-6400 (Li-COR, USA)光合仪测定旗叶净光合速率(Pn), 每个处理测3~5片叶。于10:30至11:30, 用SPAD仪测定旗叶叶绿素含量, 每个处理测定10片叶, 每片叶测5~7个点, 取平均值作为一片叶的SPAD值。
1.2.2 MDA和Pro含量 按旗叶、旗叶鞘、穗下节、颖片、籽粒5部分分别测定, 每个样品测定3次, 采用硫代巴比妥酸法[18]测定膜脂过氧化产物MDA含量; 采用磺基水杨酸法[19]测定Pro含量。
1.2.3 抗氧化酶活性 采用NBT光化还原法[18]测定超氧化物歧化酶(SOD)活性, 以20 min反应抑制NBT光化还原50%的酶量为1个酶活力单位; 采用紫外吸收法[18]测定过氧化物酶(POD)、过氧化氢酶(CAT)活性, 以每分钟A470的增加表示POD活性, 以每分钟A240的减少表示CAT活性。
1.2.4 产量和产量结构 成熟期调查穗数、穗粒数、千粒重和实际产量。每个小区调查0.5 m2穗数, 以20株平均穗粒数作为一个小区的穗粒数, 测定千粒重3次。从各小区选取2 m2样点测定实际产量, 单独收割, 脱粒称重, 籽粒含水量为13%。
用Microsoft Excel 2007整理和计算数据, 并绘图, 用SAS 8.2进行方差分析和显著性比较(LSD法)。
高温处理两小麦品种各期测定的旗叶光合速率均显著低于对照, 其光合速率平均值的降低幅度因品种而异, 河农341降低24.9%, 石家庄8号降低18.7% (图2-A)。可见, 高温处理明显降低了灌浆期旗叶的净光合速率, 但石家庄8号的旗叶光合耐热性要强于河农341。
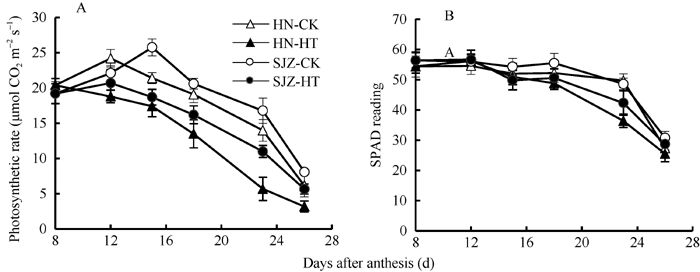 | 图2 河农341和石家庄8号旗叶净光合速率(A)和叶绿素相对含量(B)的变化(2011-2012)HN: 河农341; SJZ: 石家庄8号; HT: 高温处理; CK: 正常温度。高温处理为花后第8~22天, 处理后恢复常温生长, 取样日为花后第8、第12、第15、第18、第23和第26天。Fig. 2 Changes of net photosynthetic rate (A) and relative chlorophyll content (B) in flag leaf of Henong 341 and Shijiazhuang 8 (2011-2012)HN: Henong 341; SJZ: Shijiazhuang 8; HT: high temperature; CK: normal temperature. Wheat plants recovered normal growth temperature after the high temperature treatment from the 8th to the 22nd day after anthesis, and the sampling days were the 8th, 12th, 15th, 18th, 23rd, and 26th day after anthesis. |
高温处理下, 两品种的相对叶绿素含量均随处理时间的延长而下降, 处理第7天(花后15 d)开始明显低于对照, 在处理后第10天(花后18 d)和第15天(花后23 d), 河农341比对照分别降低6.5%和26.8%, 石家庄8号比对照分别降低8.8%和13.0% (图2-B)。可见, 高温处理会加速旗叶叶绿素衰减, 但相对而言, 石家庄8号的叶绿素衰减速率低于河农341。
对照植株器官的MDA含量以旗叶> 颖片> 旗叶鞘、穗下节> 籽粒, 高温处理后两品种旗叶、旗叶鞘、穗下节和颖片MDA含量随着处理时间的延长不断升高, 且均明显高于对照, 表明高温处理对这些器官细胞膜的伤害较为严重(图3-A1~A5)。籽粒MDA含量在灌浆期呈降低趋势, 且高温处理与对照差异很小, 显示籽粒的细胞膜稳定性强于其他器官。两品种比较, 处理和对照各器官MDA含量均是河农341高于石家庄8号; 高温处理下河农341植株旗叶、旗叶鞘、穗下节、颖片和籽粒各器官的平均MDA含量分别比对照植株相应器官高24.7%、14.2%、6.7%、11.7%和18.8%, 而石家庄8号分别比对照植株相应器官高22.2%、4.0%、6.2%、13.2%和8.3%, 说明高温胁迫导致各器官膜酯过氧化增强, 且旗叶过氧化程度明显大于非叶器官, 河农341整株受伤害程度明显大于石家庄8号。
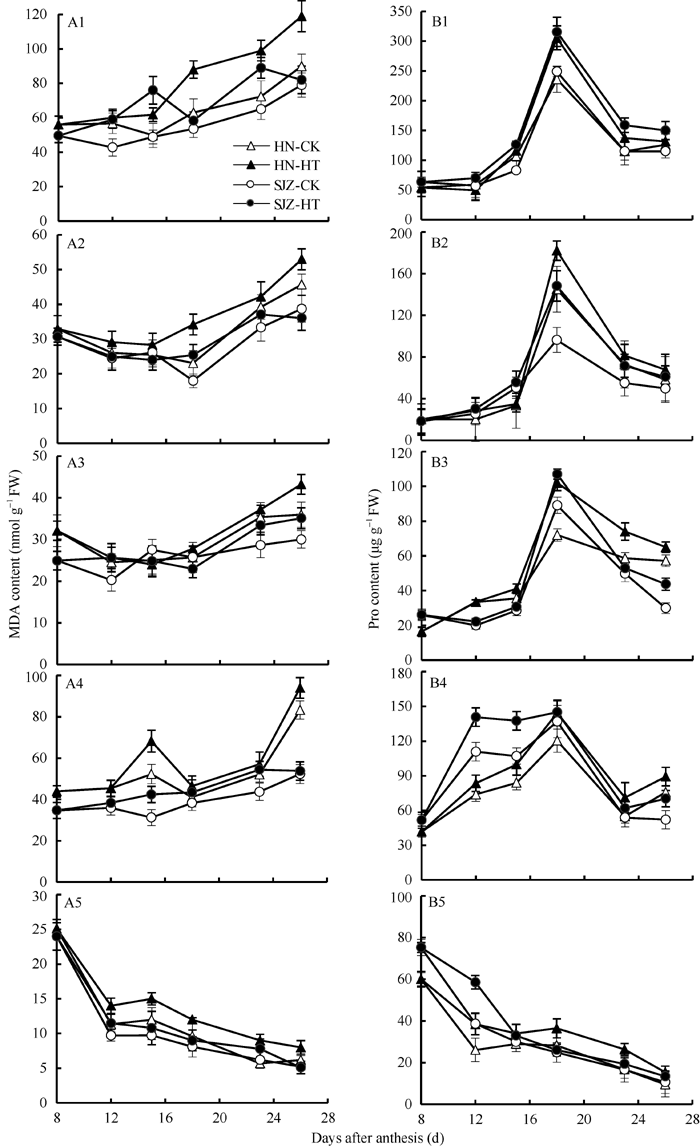 | 图3 河农341和石家庄8号高温处理和对照不同器官MDA和Pro含量的变化(2011-2012)HN: 河农341; SJZ: 石家庄8号; HT: 高温处理; CK: 正常温度。A1、B1: 旗叶; A2、B2: 叶鞘: A3、B3: 穗下节; A4、B4:颖片; A5、B5: 籽粒。高温处理为花后第8~22天, 处理后恢复常温生长, 取样日为花后第8、第12、第15、第18、第23、第26天。Fig. 3 Changes of MDA and Pro contents in different organs of Henong 341 and Shijiazhuang 8 under high and normal temperatures (2011-2012)HN: Henong 341; SJZ: Shijiazhuang 8; HT: high temperature; CK: normal temperature. A1, B1: flag leaf; A2, B2: sheath; A3, B3: peduncle; A4, B4: glume; A5, B5: grain. Wheat plants recovered normal growth temperature after the high temperature treatment from the 8th to the 22nd day after anthesis, and the sampling days were the 8th, 12th, 15th, 18th, 23rd, and 26th day after anthesis. |
在高温处理期间, 处理和对照的籽粒Pro含量呈逐渐下降趋势, 而叶与其他非叶器官Pro含量呈现先升高后降低的趋势, 最高值都出现在处理后第10天(花后18 d)。高温处理后两品种各器官Pro含量均高于对照, 说明高温诱导了Pro增加(图3-B1~B5)。从处理后各器官不同时期Pro含量平均值相对于对照增加百分比来看, 河农341表现为籽粒(24.1%)> 穗下节(21.6%)> 旗叶鞘(18.4%)> 颖片(17.6%)> 旗叶(13.8%), 石家庄8号表现为旗叶鞘(30.3%)> 旗叶(29.5%)> 颖片(18.5%)> 穗下节(16.1%) > 籽粒(15.1%)。可见, 石家庄8号高温下叶和非叶器官Pro受诱导增加程度均高于河农341。
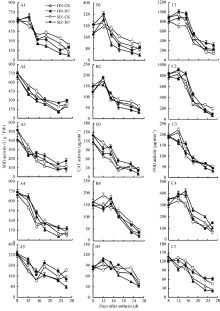
在对照条件下, 小麦灌浆期SOD活性平均值表现为旗叶> 旗叶鞘> 颖片> 穗下节间> 籽粒, 且石家庄8号各器官酶活性均高于河农341; 在高温处理下, 两品种旗叶SOD活性在处理后第4天(花后12 d)诱导上升并超过对照, 之后迅速下降, 所测灌浆期平均酶活性处理比对照显著降低, 河农341降低15.1%, 石家庄8号降低10.5%。旗叶鞘SOD活性在处理后持续下降, 处理结束后又有所回升, 整个灌浆期两品种处理的平均酶活性均低于对照, 但降低幅度均较小, 河农341为5.6%, 石家庄8号为2.5%。穗下节酶活性的变化趋势与旗叶鞘相似, 但其平均酶活性则是处理高于对照, 河农341高7.2%, 石家庄8号高8.3%。颖片SOD活性在灌浆期呈现持续下降变化, 但高温处理的平均酶活性显著高于对照, 河农341高10.8%, 石家庄8号高11.4%。籽粒SOD活性在灌浆期呈波动性下降, 但平均酶活性也以处理高于对照, 河农341高3.8%, 石家庄8号高8.7% (图4-A1~A5)。
 | 图4 河农341和石家庄8号高温处理和对照不同器官抗氧化酶活性变化(2011-2012)HN: 河农341; SJZ: 石家庄8号; HT: 高温处理; CK: 正常温度。A1~C1: 旗叶; A2~C2: 叶鞘; A3~C3: 穗下节; A4~C4: 颖片; A5~C5: 籽粒。高温处理为花后第8~22天, 处理后恢复常温生长, 取样日为花后第8、第12、第15、第18、第23和第26天。Fig. 4 Changes of antioxidant enzyme activity in different organs of Henong 341 and Shijiazhuang 8 under high and normal temperatures (2011-2012)HN: Henong 341; SJZ: Shijiazhuang 8; HT: High temperature; CK: normal temperature. A1-C1: flag leaf; A2-C2: sheath; A3-C3: peduncle; A4-C4: glume; A5-C5: grain. Wheat plants recovered normal growth temperature after the high temperature treatment from the 8th to the 22nd day after anthesis, and the sampling days were the 8th, 12th, 15th, 18th, 23rd, and 26th day after anthesis. |
在正常情况下, 小麦CAT活性表现为旗叶> 旗叶鞘、颖片> 籽粒> 穗下节间, 除颖片外, 其他器官的酶活性均是石家庄8号高于河农341; 高温处理下, 两品种旗叶片CAT活性在处理后4 d (花后12 d)迅速上升并高于对照, 但此后又迅速下降并持续低于对照。高温处理结束后, 石家庄8号酶活性有所回升但仍低于对照。各期测定的叶片酶活性平均值, 河农341处理比对照降低18.7%, 石家庄8号处理比对照降低11.2%。旗叶鞘CAT活性受高温影响的变化趋势与叶片相似, 高温处理的平均酶活性也低于对照, 降低幅度在10%以下, 且石家庄8号降幅小于河农341。穗下节CAT活性受高温影响表现出先下降后上升再下降的趋势, 平均酶活性处理高于对照, 但增幅较小, 河农341增3.6%, 石家庄8号增4.2%, 说明穗下节CAT活性相对稳定。颖片CAT酶活性在处理后也呈波动性下降, 处理的平均酶活性低于对照, 但降幅较低, 河农341为9.6%, 石家庄8号为6.4%。籽粒CAT活性在处理后的变化呈先增后降的趋势, 平均酶活性处理低于对照, 但差异很小, 河农341为2.3%, 石家庄8号为1.2% (图4-B1~B5)。
不同品种和器官POD活性及其对高温的反应有一定差异。酶活性表现为旗叶> 旗叶鞘> 颖片> 穗下节> 籽粒; 颖片POD活性以石家庄8号低于河农341, 其他器官均以石家庄8号高于河农341。在高温处理下, 两品种旗叶酶活性在高温处理前7 d (花后15 d以内)上升, 并明显高于对照, 之后迅速下降, 但仍高于对照(图4-C1~C5)。各期测定的旗叶酶活性平均值, 河农341处理比对照增加24.1%, 石家庄8号处理比对照增加14.3%。两品种旗叶鞘POD活性也在高温处理下诱导上升, 在处理前15 d (花后23 d内)均明显高于对照, 但此后低于对照, 河农341和石家庄8号的平均酶活性, 高温处理比对照分别增加4.3%和7.9%。两品种穗下节POD活性在高温处理后下降, 平均酶活性处理低于对照, 下降幅度河农341为6.3%, 石家庄8号为7.1%。两品种颖片POD活性在高温处理下的变化与叶片相似, 平均酶活性处理高于对照, 河农341高18.2%, 石家庄8号高16.5%。两品种籽粒POD活性处理与对照变化相似, 平均酶活性处理低于对照, 但差异较小, 河农341相差4.7%, 石家庄8号相差2.9%。
高温处理显著降低了两品种籽粒重量(表1), 但不同品种受影响程度不同。在2010— 2011年播期处理试验中, 晚播后期高温使河农341粒重降低15.2%, 使石家庄8号粒重降低5.1%; 2011— 2012年人工增温处理使河农341和石家庄8号千粒重分别降低37.1%和25.3%, 产量分别降低38.2%和26.1%。可见, 高温对石家庄8号粒重和产量的影响程度明显低于对河农341的影响。
| 表1 高温对石家庄8号和河农341产量及其构成因素的影响 Table 1 Effect of high temperature stress on yield and its components in Henong 341 and Shijiazhuang 8 |
大量研究显示, 高温可直接损伤光合器, 降低光合速率, 加速叶片衰老, 影响籽粒灌浆[9, 10]。本研究也表明, 灌浆期高温处理后旗叶光合速率低于对照, 且在高温处理第4天(花后12 d)或第7天(花后15 d)后显著降低, 叶绿素含量也迅速下降, 但石家庄8号光合速率和叶绿素含量下降幅度低于河农341。进一步分析发现, 高温处理下叶和非叶器官MDA含量都有所增加, 增加幅度以旗叶> 颖片> 旗叶鞘> 穗下节> 籽粒, 说明高温不仅损伤了叶片细胞膜, 也对非叶器官细胞膜造成了伤害, 但非叶器官细胞膜受损伤程度明显低于旗叶, 尤以石家庄8号表现明显。为减缓高温对原生质的损伤, 植物体内会积累Pro。本研究中, 高温处理下叶和非叶器官Pro都受诱导而含量升高, 但以非叶器官增加幅度大于旗叶, 说明非叶器官渗透调节和细胞保护能力高于叶片。
高温导致MDA含量增加后, 会增加质膜相对透性, 使植株体内氧自由基积累, 造成氧化胁迫[10, 11]。姜春明等[12]研究发现, 逆境下参与活性氧清除的SOD、CAT和POD等抗氧化酶在体内协同作用, 可增强植株清除活性氧能力, 小麦灌浆前期高温处理会诱导增强叶片中SOD、CAT酶活性, 耐热品种这两种酶活性上升更为明显。但郭天财等[7]和刘萍等[11]却发现, 高温导致了旗叶SOD、CAT、POD活性降低, 从而导致膜脂过氧化程度加剧, 衰老加快。本试验中, 小麦不同器官不同抗氧化酶活性对高温的敏感性及反应程度是不同的。旗叶、颖片和籽粒SOD活性在高温处理4 d后明显升高, 穗下节SOD活性于处理后7 d开始升高, 旗叶鞘SOD活性在处理后期增强, 说明旗叶、颖片和籽粒的SOD活性在高温处理下很快被诱导增强, 以清除体内的活性氧。但在持续高温下, 旗叶渗透调节能力较低, 细胞膜损伤较为严重, 后期SOD活性显著下降, 而非叶器官SOD活性在后期降幅较小甚至有所增强(如鞘和穗下节), 表现出相对稳定的抗活性氧能力。
SOD歧化产生的H2O2在植物体内会形成氧化力极强的羟基自由基(• OH), 由于植物细胞中缺少专一清除· OH的酶, 所以在体内积累的H2O2必须马上清除掉。植物体内清除H2O2的酶有CAT和POD。本试验结果表明, 在高温处理4 d, 两品种旗叶和籽粒CAT活性诱导增强, 旗叶鞘CAT活性石家庄8号和河农341分别于处理后4 d和7 d开始被诱导增强, 各器官CAT活性提高后有利于清除SOD作用产生的H2O2。CAT是一种诱导酶, 当植物遭受胁迫后, 植物体内H2O2浓度升高, 启动了CAT活性的表达[20], 小麦受到高温胁迫后, 短期内活性升高的SOD歧化产生更多的H2O2, 诱导CAT活性升高。处理后期上述器官的CAT活性多表现为低于对照, 说明在胁迫后期, 由于SOD活性下降, 歧化产生的H2O2含量减少, CAT活性也会下降。不过, 石家庄8号穗下节SOD和CAT活性在处理后期仍高于对照, 对于提高植株耐热性可能有重要贡献。
植物体内POD具有双重作用[21], 一方面可以催化NADH或NADPH产生O2܋, 参与活性氧的生产, 另一方面可以代谢体内积累的H2O2, 表现保护作用。本试验结果显示, 旗叶、旗叶鞘和颖片POD活性受高温诱导而明显增加, 穗下节POD活性只在处理后第7天(花后15 d)明显增强, 而籽粒POD活性总体表现低于对照。从各器官酶活性来看, 在高温处理初期POD可能表现保护效应, 协同CAT共同代谢旗叶、旗叶鞘和颖片中的H2O2, 减轻高温胁迫的氧化伤害, 而在高温处理后期POD可能表现伤害效应, 加速旗叶和颖片的衰老, 在穗下节和籽粒中, SOD歧化产生的H2O2可能主要靠CAT清除, 是否如此, 还需进一步研究。小麦抗氧化胁迫能力与体内SOD、CAT和POD的活性及其协同作用密切相关[22]。根据本试验结果, 高温下石家庄8号的SOD、CAT和POD活性增加幅度均高于河农341, 说明石家庄8号抗氧化胁迫综合能力强于河农341。由于小麦耐热性存在遗传变异[23], 对这两个品种不同器官抗氧化能力差异的遗传生理机制有待进一步研究。
高温胁迫下, 小麦非叶器官相对于叶片表现出膜质过氧化程度较低、脯氨酸含量增幅较高和抗氧化酶活性较稳定的特征。受高温胁迫, 石家庄8号旗叶光合的降幅小于河农341, 且叶和非叶器官细胞膜稳定性、抗氧化酶活性及其增加幅度均高于河农341, 这可能是石家庄8号耐热性强于河农341、最终产量下降幅度低于河农341的重要原因。鉴于非叶器官具有较强的耐热性, 我们认为, 为增强小麦对气候变化的适应性, 在小麦抗逆育种工作中应重视对非叶光合器官形态与功能特性的选择和评价, 在小麦高产栽培中应重视并优化调整非叶器官在群体结构中的配置模式, 以充分利用和发挥其耐逆机能优势。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|