{kind=link}
Glu-1位点缺失对小麦麦谷蛋白聚合体粒度分布及面团特性的影响
[张平平1, *  , 马鸿翔
, 马鸿翔1 , 姚金保1 , Joseph M. AWIKA2 ]
, 马鸿翔|
|


以 Glu-1位点正常和部分缺失的小麦品系为材料, 探讨HMW-GS和LMW-GS组成与谷蛋白聚合体粒度分布和面团特性的关系, 为利用HMW-GS缺失系改良小麦品质提供理论依据。在20个供试硬白冬麦品系中, 1个品系为 Glu-A1位点缺失, 5个品系为 Glu-D1缺失, 3个品系为 Glu-A1和 Glu-D1双缺失。所有品系的蛋白质含量皆较高(13.39%~14.12%), 品系间无显著差异, 缺失系与非缺失系间也无显著差异。 Glu-1位点缺失显著降低了高分子量谷蛋白/低分子量谷蛋白比(HMW/LMW)、不溶性谷蛋白大聚体的含量和百分比。谷蛋白/醇溶蛋白比(GLU/GLI)在基因型间变幅较小, 且在缺失系和非缺失系间无显著差异。 Glu-1位点缺失显著降低了面团弹性, 但显著提高了面团的延展性。部分 Glu-1位点缺失系仍具有较高的面团强度和突出的延展性, 谷蛋白聚合体粒度分布和面团特性受谷蛋白亚基组成和表达量的共同影响。研究结果表明, 利用 Glu-1位点亚基缺失可能是改善面筋延展性, 提高食品加工品质的方法之一。
The relationship between the HMW-GS and LMW-GS components and the size distribution of glutenin polymeric protein and dough properties were studied using Glu-1 normal and deletion lines, which provides a foundation for investigating the potential of Glu-1 deletion line in quality improvement of wheat. In the 20 hard white winter wheat lines tested, one was Glu-A1deletion line, five were Glu-D1deletion lines, and three were Glu-A1 and Glu-D1double-deletion lines. Flour protein contents of the lines tested ranged from 13.39% to 14.12% with no significant difference between each other and between deletion and non-deletion groups. In Glu-1deletion lines, the high molecular weight glutenin subunits/low molecular weight glutenin subunits ratio (HMW/LMW), content of SDS unextractable polymeric protein (UPP), and percentage of UPP (%UPP) were significantly lower than those in non-deletion lines. However, the glutenin/gliadin ratio (GLU/GLI) was similar in all genotypes, with no significant difference between deletion and non-deletion groups. Deletion at Glu-1 resulted in a significant decrease of dough elasticity and a significant increase of dough extensibility. Several Glu-1 deletion lines were characterized with medium gluten strength and excellent extensibility, suggesting that the size distribution of glutenin polymeric protein and dough properties were affected by both allelic composition and quantity of glutenin subunits. These results indicate that Glu-1 deletion lines can be used to improve dough extensibility and processing quality of common wheat.
蛋白质约占小麦籽粒重量的8%~20%, 其中贮藏蛋白约为总量的85%, 由谷蛋白(glutenin, GLU)和醇溶蛋白(gliadin, GLI)组成。谷蛋白则由高分子量谷蛋白亚基(high molecular weight glutenin subunit, HMW-GS)和低分子量谷蛋白亚基(low molecular weight glutenin subunit, LMW-GS)组成。因HMW-GS易于分离, 与加工品质密切相关, 前人以不同的研究材料和品质指标建立了相应的HMW-GS评分方法[1, 2, 3]。以传统的杂交育种和现代生物工程技术操纵小麦的HMW-GS组成已成为优质小麦育种的重要途径[4, 5, 6]。
HMW-GS由Glu-A1、Glu-B1和Glu-D1 3个Glu-1位点编码, 每位点编码x型和y型2个亚基; LMW-GS则由Glu-A3、Glu-B3和Glu-D3 3个Glu-3位点编码, 较HMW-GS具有更丰富的多态性[7, 8]。HMW-GS和LMW-GS通过二硫键形成大小不同的聚合体(glutenin polymer, GP)作为面筋网络的结构单位和功能单元[9]。利用SDS缓冲液中可将谷蛋白聚合体分离为可溶性(SDS-extractable polymeric protein, EPP)和不溶性(或谷蛋白大聚体, SDS- unextractable polymeric protein, UPP)两种聚合体, 前者粒度较小后者粒度较大, 两者的含量和比例影响面筋的弹性和延展性, 以影响面筋的弹性为主[10, 11]。因此, Glu-1位点和Glu-3位点的组成、含量及互作通过改变聚合体的粒度分布进而影响面团特性和加工品质[11, 12, 13, 14, 15]。
突变体和近等基因系是探讨谷蛋白亚基功能的重要方法, Larroque等[16]利用突变体研究发现, Glu-1位点缺失显著降低了UPP含量, 但与面筋质量和加工品质的关系未作探讨。张莉丽等[5]则利用近等基因系探明了Glu-A1位点1亚基表达和缺失间存在显著的面筋品质差异, 但未探讨野生型中不存在的Glu-B1和Glu-D1 位点缺失效应。武茹等[17]通过Glu-A1和Glu-D1双缺失位点的杂交转育, 显著降低了弱筋小麦的沉降值和水溶剂保持力, 但并未详细探讨其对贮藏蛋白和面筋质量的影响。Jondiko等[18]则进一步研究了HMW-GS缺失与墨西哥卷饼加工品质的关系, 并提出HMW-GS表达位点数量的操控可能是特殊食品加工品质提高的有效途径, 但未探讨与HMW-GS和LMW-GS组成的关系。目前多数研究者仍集中于HMW-GS缺失基因型的创制和发掘[19, 20, 21, 22], 以及缺失机制的探讨[20], 对其与面筋蛋白功能单元以及面团特性的影响鲜见报道。本文旨在利用HMW-GS正常表达和Glu-1部分位点缺失的高蛋白含量品系为材料, 探讨HMW-GS和LMW-GS组成与谷蛋白聚合体粒度分布和面团特性的关系, 为利用HMW-GS缺失改良小麦品质提供理论依据。
试验材料为20份具有不同HMW-GS和LMW- GS组成的硬白冬麦(表1), 由美国Texas A& M University的Cereal Quality Lab提供。其中L1~L9、L11和L14的HMW-GS正常表达, L10、L12~L13、L15~L20则利用HMW-GS缺失突变体Ogallala及衍生系通过常规杂交育种途径选育, 试材间的遗传背景不同。试验材料于2011— 2012年度种植于美国德克萨斯北部的McGregor和Bushland。田间试验采用随机区组设计, 3次重复, 收获后3次重复等量混匀。田间管理按一般品种比较试验。
| 表1 参试品系的高低分子量麦谷蛋白亚基组成 Table 1 High-molecular-weight glutenin subunits composition of wheat lines |
使用单籽粒谷物分析仪(Perten Instruments North America Inc., Springfield, IL, USA), 按AACC 55-31.01[23]方法测定籽粒硬度, 硬度范围为50~75。使用Brabender Quadrumat Senior Laboratory 按照标准程序制粉, 出粉率65%~72%。使用Perten DA7000近红外分析仪(Perten Instruments North America Inc., Springfield, IL, USA), 按AACC 39-11.01[23]方法测定蛋白质含量。按AACC 54-40.02利用10 g微量揉面仪测定面团揉面峰值时间(National Manufacturing, Lincoln, NE)。利用物性仪(Model TA-XT2i, Texture Technologies Corp., Scarsdale, NY/Stable Micro Systems, Godalming, Surrey, UK)拉升测试模块测定面团拉伸最大抗阻和延展性。
参考Singh等[24]和Liu等[25, 26]的SDS-PAGE法分离高低分子量麦谷蛋白亚基(HMW-GS)。通过分离总谷蛋白鉴定HMW-GS和LMW-GS的Glu-A3; 通过分离醇溶蛋白, 并根据醇溶蛋白与Glu-B3的连锁关系鉴定LMW-GS的Glu-B3。采用经修改的反相高效液相色谱法(reversed-phase high-performance liquid chromatography, RP-HPLC)分离和量化HMW- GS和LMW-GS, 并计算高、低分子量谷蛋白含量的比值(HMW/LMW) [27]。参考Larroque等[28]的凝胶色谱法(size-exclusion high-performance liquid chromatography, SE-HPLC)测定谷蛋白/醇溶蛋白含量比(GLU/GLI)、SDS-可溶性谷蛋白聚合体(SDS- extractable glutenin polymeric protein, EPP)和SDS-不溶性谷蛋白聚合体(SDS-unextractable glutenin polymeric protein, UPP)的含量, 同时计算UPP占谷蛋白聚合体总量的百分比, %UPP = UPP / (EPP + UPP) × 100。
用Statistical Analysis System 9.0统计分析软件对供试品种的蛋白质含量、谷蛋白聚合体粒度分布参数和面团品质参数进行基本统计量和简单相关性分析。以试点和基因型为变异因素进行方差分析(PROC GLM), 并在基因型间进行显著性比较。将HMW-GS缺失系和非缺失系聚类后对两类品系的相关参数进行显著性比较。
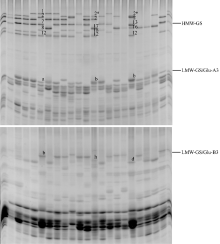
20个参试品系的HMW-GS和LMW-GS变异丰富, Glu-A1位点有3种亚基, Glu-B1位点有5种, Glu-D1位点有3种, Glu-A3位点有3种, Glu-B3位点有4种(表1)。11个品系为HMW-GS非缺失品系, 9个品系为HMW-GS部分缺失品系。其中L19为Glu-A1单位点缺失, L10、L12、L13、L15和L20的Glu-D1单位点缺失, L16、L17和L18则为Glu-A1位点和Glu-D1位点双缺失(图1)。
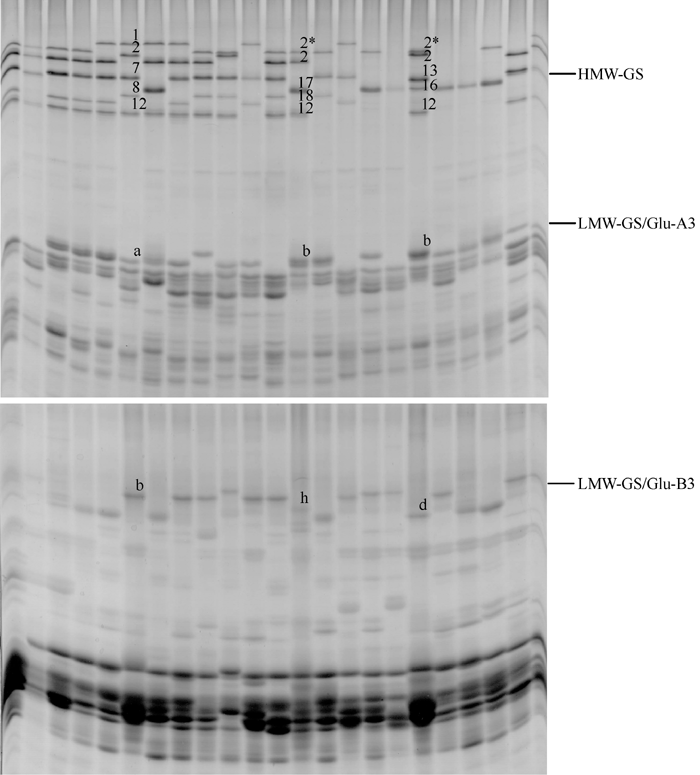 | 图1 20个参试小麦品系的高低分子量麦谷蛋白亚基聚丙烯酰胺凝胶电泳图谱Fig. 1 SDS-PAGE profiles of HMW-GS and LMW-GS in 20 wheat lines tested |
方差分析表明, 除蛋白质含量外, GLU/GLI、HMW/LMW和谷蛋白大聚体粒度分布参数在基因型间存在显著差异; 除GLU/GLI外, 环境间各参数也存在显著差异。不同品系间的蛋白质含量差异较小, 变幅仅为0.76% (表2)。尽管L12为Glu-D1位点缺失, 但蛋白质含量最高为14.12%; 相反, L4和L14的Glu-1位点虽正常表达, 但蛋白质含量表现最低, 为13.39%和13.36%, 这与HMW-GS在贮藏蛋白总量中含量较低有关。
| 表2 高分子量麦谷蛋白亚基缺失对蛋白质含量和谷蛋白大聚体粒度分布的影响 Table 2 Effects of HMW-GS deletion on protein content and the size distribution of polymeric protein |
品系间的GLU/GLI变异范围为0.50~0.76, 最小值和最大值均为HMW-GS正常表达的品系, 与Glu-1位点缺失与否无直接关系。HMW-GS缺失显然降低了其在谷蛋白中的相对含量, HMW/LMW总体表现低于非缺失系。其中, 缺失品系的HMW/ LMW变异范围为0.22~0.33, 非缺失系的HMW/ LMW变异范围为0.32~0.56。缺失品系L18和L19的HMW/LMW值(0.33)略高于非缺失系L8和L9 (0.32), 表明HMW/LMW受Glu-1位点表达数量和HMW-GS表达量的共同影响。
缺失系和非缺失系的EPP含量平均值分别为7.71AU和7.04AU, 差异显著 (表2), 但两组中个别基因型间并无明显的消长关系。相反, 缺失系的UPP含量和%UPP则显著低于非缺失系, 其中缺失系和非缺失系的UPP含量平均值分别为3.61AU和5.45AU, 表明HMW-GS缺失显著降低其含量(表2)。个别HMW-GS缺失品系的UPP含量高于非缺失品系, 如缺失系L10的UPP含量为4.42AU, 高于非缺失系L5的含量, 可能是由于L5的Glu-A3位点为e (缺失), 对面筋强度的贡献效应最低, 不利于谷蛋白大聚体的形成[13]。又如缺失系L13, 虽Glu-D1位点缺失且Glu-A3位点为e, 但UPP含量仍高于部分非缺失品系。%UPP为相对值, 与UPP存在极显著的正相关性(r = 0.64, P= 0.001), 因此%UPP也显示出与UPP相似的变化规律, 即大多数非缺失系的%UPP显著高于缺失系。个别非缺失品系如L5、L8、L9和L14的%UPP略低于缺失系或与缺失系相似, 可能与HMW-GS和LMW-GS的组成、表达量及互作有关[12, 13, 14, 15].
缺失系与非缺失系的籽粒硬度平均值分别为59.4和64.0, 变异范围分别为46.2~69.2和52.4~75.2, 两者间无显著差异(表3), 但面团特性显著不同, 表明硬度和面筋质量是相对独立的两个性状。揉面仪峰值时间与拉伸最大抗阻高度正相关(r = 0.98, P= 0.001), 因此HMW-GS缺失与否对两者具有相似的影响。缺失系的峰值时间和最大抗阻显著低于非缺失系, 而延展性显著高于非缺失。缺失系的峰值时间和拉伸最大抗阻的变异范围为1.30~2.94 min和0.06~0.23 N, 非缺失系相应的变异范围为2.70~ 5.20 min和0.16~0.63 N。如L10与L6相比, 1/7+9/ null/a/b和1/7+9/5+10/a/b, 峰值时间分别为1.70 min和5.20 min。同时, 缺失系L17和L18的HMW-GS虽为Glu-A1位点和Glu-D1位点双缺失, 但面筋强度仍较高, 而L5、L8、L9和L14虽HMW-GS正常表达但面筋强度则相对较低, 这与谷蛋白大聚体粒度分布参数中%UPP的变化规律相似(表2)。表明HMW-GS缺失可显著降低面筋强度, 但仍然受到谷蛋白亚基质量组成的影响, 而延展性则表现相反, 缺失系和非缺失系的变异范围分别为102.3~143.0 mm和71.1~126.5 mm, HMW-GS缺失明显改善了面筋的延展性。
| 表3 高分子量麦谷蛋白亚基缺失对面团特性的影响 Table 3 Effects of HMW-GS deletion on kernel hardness and dough properties |
HMW/LMW、EPP、UPP和%UPP与面团特性高度相关, 且与面筋强度和延展性的关系相反(表4)。HMW/LMW、UPP和%UPP的提高可增加面筋强度, 而EPP的提高则可改善面筋的延展性, 其中以%UPP对面筋质量的影响最显著, 与峰值时间、最大抗阻及延展性的相关系数分别为0.95、0.92和-0.82, 达极显著水平。表明谷蛋白亚基的组成通过改变大聚体的粒度分布进而改变面筋强度。
| 表4 蛋白质含量及贮藏蛋白特性参数与籽粒硬度和面团特性的相关系数 Table 4 Correlation coefficients between flour protein content, gluten protein parameters and dough properties |
尽管HMW-GS等位变异在面筋质量和加工品质中的重要性已经得到充分验证[1, 2, 3, 8, 29], 但由于HMW-GS间, 以及与LMW-GS的互作, 使得HMW- GS的重要性在不同基因型中表现不同[8, 13, 30]。进一步的研究又显示, 除了亚基间的加性和互作效应外, 高低分子量谷蛋白亚基的表达量又进一步影响既定亚基组成下的面筋质量和加工品质[14, 31]。基因沉默是研究基因功能的重要方法, 通过部分或全部HMW-GS的缺失可探讨单个亚基或单个位点, 以及亚基或位点间互作对加工品质的贡献。因此, 众多研究者开展了小麦谷蛋白亚基缺失突变体的创制和研究[5, 16, 17], 但亚基缺失对谷蛋白大聚体功能单元和和加工品质的关系仍需明确。其中Larroque等[16]利用突变构建的近等基因系为材料表明, Glu-1缺失位点增多, UPP含量显著降低; HMW-GS组合为1/17+18/5+10的品系%UPP达49.5%, 而Glu-1位点全部缺失的品系仅为11.7%, 其中以Glu-B1和Glu-D1位点的贡献最大。这与Glu-B1x和Glu-D1x亚基在5个正常表达HMW-GS中含量较高有关[14, 31]。本研究所使用的材料没有Glu-1位点全部缺失的品系且并非近等基因系, 因此不能准确探知Glu-1位点缺失数量或HMW-GS缺失数量对谷蛋白大聚体粒度分布的影响。正如文中多数HMW-GS缺失系不同程度降低了不溶性谷蛋白大聚体的含量和面筋强度, 但少数缺失系仍表现较高, 相同Glu-1位点缺失的品系间大聚体粒度分布参数和面筋质量仍存在较大差异。如缺失系L16、L17和L18尽管同时缺失Glu-A1和Glu-D1, 但此3个品系的%UPP和面筋强度差异较大, %UPP和面筋强度较缺失1个位点的品系相似或较高。与UPP相比, EPP中的HMW/LMW比值较低[31, 32], 因此其受HMW-GS缺失的影响相对较小。同时, 尽管EPP在缺失系和非缺失系类型间存在显著差异, 但两种类型间单个基因型的比较并无明显的规律。综上可进一步证实谷蛋白亚基间的互作、谷蛋白亚基含量、HMW-GS缺失共同影响了谷蛋白大聚体粒度分布。研究表明, Glu-1位点对谷蛋白大聚体粒度分布的影响还受到蛋白质含量的影响[15, 34, 35], 但本研究各基因型的蛋白质含量变异范围较小(13.39%~14.12%), 全部品系间、缺失系与非
缺失系间均无显著差异, 其原因不仅与遗传背景有关, 亦与HMW-GS在贮藏蛋白总量中所占比例较小有关。因此, 应选育谷蛋白组成和蛋白质含量水平均不同的近等基因系, 以进一步探明HMW-GS效应。
HMW-GS、LMW-GS和GLI分别占小麦贮藏蛋白总量的约10%、40%和50%, 三者的数量和质量变异影响面团的弹性、延伸性以及加工适用性[7]。HMW- GS的含量虽很低但对加工品质起决定性作用[1, 2, 3]。本研究亦证实HMW/LMW与面筋质量显著相关, 但GLU/GLI与面筋质量无显著相关性, 这与本研究所选用的材料有关。在本研究中, 部分材料谷蛋白总量较高, 但HMW-GS缺失或其组成较差, 最终导致品质降低。尽管GLU/GLI对面筋质量的贡献较小, 但谷蛋白组成及相对含量与面筋质量高度相关, 进一步表明谷蛋白质量是影响面筋质量最重要的因素, 尤其HMW/LMW和%UPP两个相对含量参数对面筋质量具有决定性作用[14, 36]。适当的面团弹性和良好的延展性是各类面粉制品的共同需求。延展性与蛋白含量、GLU/GLI和HMW/LMW呈负相关, 因此面筋强度和延展性在强筋小麦的改良中具有矛盾性。我们曾对北方冬麦区大面积推广的优质冬小麦品种和常用亲本进行分析, 多数材料蛋白质含量较高且含有优质亚基, 但延展性和加工品质不及国外对照品种, 如藁城8901拉伸仪最大抗阻达540 BU, 但延伸性不足170 mm [14]。弱筋小麦同样存在延展性不足的问题, 如对宁麦9号衍生系的面团吹泡特性分析后发现, 多数品系的延展性不足70 mm [37], 较优品系的延展性大多也不足90 mm [38], 与大于106 mm的优质标准相比, 仍待提高[39]。相比面团强度, 延展性更需要谷蛋白亚基、谷蛋白与醇溶蛋白在质量和数量上的协调[9]。总的来看, HMW-GS位点缺失对面筋强度和延展性具有相反的作用, 但在具体品系中的表现不同。部分品系由于HMW-GS缺失而弹性大幅下降, 延展性大幅提高; 但部分品系缺失HMW- GS后弹性适中且延展性大幅提高, 如缺失系L17、L18、L19和L20。通过传统的杂交育种方法改良延展性较为困难, HMW-GS亚基构成位点简单, 通过品种选择和亚基缺失修饰可显著改善面筋延展性。Jondiko等[18]对HMW-GS缺失系的研究表明, 一个HMW-GS组成为2* /17+null/5+null和蛋白含量为13.7%的品系具有中等面筋强度和突出的延展性, 极显著地提高了墨西哥卷饼的直径和货架寿命, 暗示HWM-GS缺失可能是进一步改良食品加工品质的途径之一。LMW-GS等位变异对面筋质量的重要影响则体现在与HMW-GS的互作及对总含量的贡献[13, 14, 25, 26, 36]。
本研究选用材料中的HMW-GS组成及缺失类型有限且背景不同, 亦未对HMW-GS单个亚基进行定量分析。但由于Glu-1位点是面筋质量的主要影响因子, 可以初步探讨不同位点的缺失效应。
无论Glu-1位点缺失与否, 参试品系具有相似的蛋白质含量, 谷蛋白聚合体粒度分布和面筋质量受谷蛋白亚基组成和表达量的影响。HMW-GS缺失显著降低UPP、%UPP和HMW/LMW值及面团弹性, 但提高面团延展性。HMW-GS缺失可能是改善面团延展性、提高食品加工品质的方法之一。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|