
{kind=link}
{kind=link}
{kind=link}
{kind=link}
拟南芥下胚轴向光弯曲 P2SA2基因的克隆与功能鉴定
[赵青平, 赵翔, 慕世超, 肖慧丽, 张骁*  ]
]
]
|
|


向光素PHOT1介导较宽范围蓝光诱导的下胚轴向光弯曲, 而向光素PHOT2仅在强蓝光下起作用。强蓝光下, PHOT1和PHOT2介导拟南芥下胚轴向光弯曲的功能冗余性, 限制了人们对PHOT2功能的研究。为此, 以拟南芥 phot1突变体为材料, 避开 PHOT1基因的干扰, 通过EMS诱变筛选拟南芥下胚轴向光不弯曲突变体, 成功克隆到1个基因, 命名为 P2SA2 ( phototropin 2 signaling associated 2), 该基因被证明是 NPH3的等位基因。 P2SA2基因的突变可导致拟南芥缺失强蓝光诱导的下胚轴向光弯曲反应。在 p2sa2突变体背景下, P2SA2基因超表达可恢复强蓝光诱导的拟南芥下胚轴向光弯曲。该结果将为强蓝光下PHOT2下游基因的筛选、功能鉴定和揭开PHOT2调节强蓝光诱导的下胚轴弯曲的机制提供理论基础。
PHOT1 functions at both low- and high-intensity blue light to mediate phototropic responses, but PHOT2 functions only at high-intensity blue light. The functional redundancy of PHOT1 and PHOT2 on high-intensity blue light-induced phototropic curvature of hypocotyls in Arabidopsis thaliana, restricts the understanding of the mechanism of PHOT2 signal transduction. Therefore, in order to avoid the interference of PHOT1, Arabidopsis phot1 mutant was selected as material for screening high blue light insensitive mutants by the EMS mutation, and we successfully screened and cloned the gene P2SA2 ( phototropin 2 signaling associated 2). The gene P2SA2turned out to be the allelic of NPH3(Nonphototropic hypocotyl 3). The mutation of gene P2SA2 could result in that Arabidopsis thaliana lost the phototropism to unilateral high intensity blue light. Transgenic plants of p2sa2 35S::P2SA2 restored hypocotyl phototropism to high intensity blue light. These findings will open new perspectives about the screening and functional identification of PHOT2 downstream genes in response to high blue light, and provide the theoretical basis to uncover hypocotyl bending mechanism regulated by PHOT2.
植物向光性是植物对光产生的最普遍反应, 即在单侧光照射时, 植物体部分向光发生弯曲生长。已有研究表明, 向光性由多个光受体系统控制, 尤其是蓝光受体向光素。拟南芥蓝光受体向光素(PHOT1和PHOT2)的C末端含有Ser/Thr蛋白激酶区域[1], N端含有与FMN结合对光照、氧气及电压差敏感的2个LOV (light, oxygen, voltage)区, 调节其C末端激酶活性[2]。蓝光激发引起一个可逆的光循环, FMN和LOV区内保守的半胱氨酸之间形成共价结合, 诱导蛋白构象变化, 激活C端激酶区, 引起受体自磷酸化[3], 从而引起相应生理反应, 如植物向光性[4], 气孔运动[5]、叶绿体移动反应[6]以及叶片伸展[7]等, 优化植物在极弱或极强光逆境下的生长[8, 9], 但其具体的分子机制并不清楚。
PHOT1介导弱蓝光(0.01~1.00 µ mol m-2 s-1)和强蓝光( > 1.00 µ mol m-2 s-1)诱导的下胚轴向光弯曲, 而PHOT2仅在强蓝光( > 1.00 µ mol m-2 s-1)下起作用[4, 10]。目前, 关于PHOT1介导弱蓝光引起的下胚轴弯曲研究较为明确, 即PHOT1感受弱蓝光后, 与其下游的信号蛋白NPH3[11]、RPT2[12]和PKS1[13]相互作用, 调控生长素输出载体PIN1和输入载体AUX1[14, 15]的活性及定位, 引起生长素在下胚轴中不对称分布, 导致植物向光弯曲。虽已证明NPH3和RPT2也参与PHOT2介导强蓝光诱导的下胚轴向光弯曲调节[12, 16], 但并未检测到RPT2与PHOT2体内互作[12, 13]。近年研究发现, 编码Ser/Thr蛋白磷酸酶2A (PP2A)A1亚基的RCN1与PHOT2蛋白体外互作, 双突变体phot1-5rcn1-1在蓝光的照射下, 表现增强拟南芥下胚轴向光弯曲[17]。因此, 进行强蓝光下PHOT2下游基因的筛选与功能鉴定, 将为揭开PHOT2调节强蓝光诱导下胚轴弯曲的机制提供基础。
文中以phot1突变体为材料, 利用EMS (甲基磺酸乙酯)诱变筛选下胚轴强蓝光不弯曲突变体, 以期获得PHOT2下游信号分子或PHOT1和PHOT2信号的中心调节子。目前, 成功克隆到1个基因, 命名为P2SA2 (phototropin 2 signaling associated 2), 该基因被证明是NPH3的等位基因。P2SA2基因的突变, 可导致拟南芥缺失强蓝光诱导的下胚轴向光弯曲反应。构建P2SA2基因的超表达载体获得p2sa2 35S:: P2SA2植株, 发现其恢复了强蓝光诱导的拟南芥下胚轴向光弯曲反应。通过对P2SA2基因的研究, 将为揭示受PHOT1和PHOT2调节的强蓝光诱导下胚轴弯曲的差异机制提供理论依据。
拟南芥(Arabidopsis thaliana L.)生态型除用于图位克隆的Landsberg (Ler)为野生型外, 其余都是Columbia-0生态型(gl1), 用作野生型对照, 拟南芥突变体种子phot1 (phot1-5)、phot2(phot2-1)和phot1phot2(phot1-5 phot2-1)由Ken-ichiro Shimazaki (日本九州大学)惠赠[6, 18], 突变体nph3-6由Christian Fankhauser (瑞士日内瓦大学)惠赠[11]。试验以拟南芥phot1突变体为材料参考赵翔等[19]的方法进行诱变。
将拟南芥M2代种子点种于0.6% MS培养基, 4℃低温处理3 d, 待下胚轴长度约1 cm时, 移入0.8% MS培养基, 将培养皿垂直放置, 进行单侧强蓝光(100 μ mol m-2 s-1)处理。处理12 h后观察M2代幼苗下胚轴向光弯曲情况, 筛选出下胚轴向光不弯曲的疑似突变体移出进行培养, 待单株收获种子后进行遗传和生理分析。突变体遗传分析, 以稳定遗传的疑似突变体为母本, 分别与父本phot1和WT (Landsberg生态型)回交和杂交得到F1代, F1代自交后产生群体F2。把F2点种在含有0.6%琼脂的MS培养基上, 4℃低温处理3 d后暗处理4 d, 将黄化幼苗移入0.8% MS培养基, 进行强蓝光(100 μ mol m-2 s-1)单侧处理。观察下胚轴向光弯曲与不弯曲的比例, 以确定其遗传特性。
参照Zhao等[20]的方法测量下胚轴向光弯曲度, 将黄化4 d生长约5~8 mm的拟南芥幼苗用镊子小心移入0.8% MS培养基, 整齐排列成两行, 使下胚轴与根部都紧贴培养基表面, 垂直放于23℃暗室, 照水平单侧蓝光12 h。用数码相机照相, 在电脑上用电子软件E-尺测量弯曲度数。试验重复3~5次, 统计其平均值。用t检验进行差异显著性分析。
提取野生型拟南芥叶片RNA并将其进行反转录合成cDNA, PCR扩增P2SA2 CDS片段全长并回收目的片段, 将目的基因与相关载体空质粒同时进行双酶切, 连接产物转化大肠杆菌, 37℃过夜培养后, 选取生长状态良好的菌落, PCR鉴定是否为阳性菌落, 阳性菌落提取的质粒再经过双酶切和质粒PCR验证。将上述阳性质粒转化农杆菌GV-3101, 鉴定后保存阳性菌落菌种。选取生长旺盛, 开花较多的拟南芥进行转化, 转化前剪掉长出的果荚。将其花浸于菌液(含有目的片段质粒的农杆菌GV-3101)中保持30 s, 然后拿出来标记好, 平放在转苗用的铁托盘中。浸染完所有植株后, 用保鲜膜保持其湿润状态, 并将其于避光处放置24 h。随后将侵染过的材料取出正常培养。为增加农杆菌的转化效率, 可在第1次转化1周新的花蕾长出后, 进行第2次侵染。待收获种子后使用潮霉素(Hyg)筛选潜在的转基因植株。
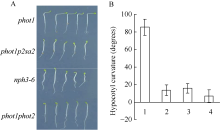
考虑到PHOT1和PHOT2功能冗余介导强蓝光诱导的拟南芥下胚轴向光弯曲生长, 为避开PHOT1基因的干扰, 寻找PHOT2下游信号分子, 我们以phot1突变体(遗传背景为Columbia生态型)为材料, 利用0.3% EMS (甲基磺酸乙酯)诱变, 将诱变的种子种下, 自交一代后得到M2代。从大约8万粒M2代种子中初步筛选出3株, 在100 µ mol m-2 s-1单侧蓝光照射12 h下胚轴弯曲情况与phot1存在明显不同的突变体(图1-A)。随后将其单株收种, 再次进行表型验证, 如图1-B和图1-C所示, 初筛的3株疑似突变体依然表现出对强蓝光刺激黄化苗下胚轴向光不弯曲的表型(WT为Columbia生态型)。基于PHOT1基因已经突变, 新突变体中的突变基因应该是PHOT2或位于PHOT2下游参与调节拟南芥下胚轴向光弯曲反应的某个基因, 又由于该突变体是在phot1突变体背景下筛选获得, 因此命名为phot1p2sa2 (phototropin2 signaling associated 2)。
 | 图1 PHOT2介导拟南芥下胚轴向光弯曲下游调节子筛选 A: 从EMS诱变phot1突变体M2代群体中筛选强蓝光不敏感突变体; 红框: 疑似突变体phot1p2sa1; 绿框: 疑似突变体phot1p2sa2; 黄框: 疑似突变体phot1p2sa3; 箭头: 照光方向、强度和处理时间; B: 100 µ mol m-2 s-1蓝光单侧处理12 h后不同基因型拟南芥下胚轴向光弯曲表型; C: 不同基因型拟南芥下胚轴向光弯曲度统计。图中每个数据分别来自3次独立重复试验, 大约19~24颗苗的平均值± 标准误。其中1: WT; 2: phot1; 3: phot1p2sa1; 4:phot1p2sa2; 5: phot1p2sa3。Fig. 1 Isolation of regulator involved in PHOT2-mediated phototropism of hypocotyls inArabidopsis A: HBL insensitive mutants fromArabidopsis M2 mutagenesis population of phot1 mutants and selfing M1; red box: phot1p2sa1; green box: phot1p2sa2; yellow box: phot1p2sa3; Arrows: direction of light irradiation, fluence rate and treated time; B: typical seedling phototropic curvature in indicated lines at 100 µ mol m-2 s-1 blue light for 12 hours; C: hypocotyl curvatures were measured in indicated lines. 1: WT; 2:phot1; 3:phot1p2sa1; 4:phot1p2sa2; 5:phot1p2sa3. Values are the means ± SD (n=19-24). |
首先将phot1p2sa2突变体与phot1回交, 发现F1代下胚轴向光均发生弯曲, 而F2出现下胚轴向光弯曲与不弯曲表型分离为比3∶ 1。将phot1p2sa2突变体与拟南芥野生型(Col生态型)杂交, 发现与phot1突变体回交结果相似, F1代下胚轴均发生向光弯曲反应, F2表型分离比同样出现3∶ 1 (表1), 证明phot1p2sa2突变体的表型为单基因隐性突变所致, PHOT1基因突变与否并不影响对p2sa2突变位点的克隆。同时将突变体phot1p2sa2与野生型(Landsberg生态型)杂交, 其F1代下胚轴均发生向光弯曲, F2分别出现下胚轴弯曲和不弯曲表型分离比依然为3∶ 1 (表1), 再次证明phot1p2sa2突变体表型是单基因隐性突变。同时为其突变位点的克隆提供材料。
| 表1 突变体杂交遗传分析 Table 1 Genetic analysis of mutants |
选取拟南芥5条染色体中每条染色体上、中、下游分子标记, 共15个, 对phot1p2sa2突变体(Col生态型)与野生型(Ler生态型)杂交F2代下胚轴向光不弯曲的幼苗(接近100棵)提取DNA, 进行PCR扩增, 电泳检测统计每个分子标记的重组率, 如表2所示, 发现MQB2这个BAC的重组率最低, 为6.32%, 可以判断p2sa2的突变位点, 可能位于拟南芥第5染色体的下部。
| 表2 粗定位引物及重组计算 Table 2 Primers for locations and recombinant frequency |
为此, 在第5染色体MQB2的BAC前后, 选定了6个有效的分子Marker, 进行细定位。结果如表3, MSJ1这个BAC上的重组率最低, 为0.23%, 最终将p2sa2突变基因定位到MSJ1的BAC上。
| 表3 细定位引物及重组计算 Table 3 Primers for fine positioning and recombinant frequency |
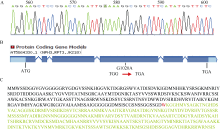
MSJ1这个BAC全长88.41 kb, 共有17个基因, 右端与MGI19有8.53 kb重叠, 左端与MPA24有7.56 kb重叠, 将MSJ1以及与MGI19、MPA24相重叠的基因, 根据拟南芥网站(http://arabidopsis.org/)上报道基因信息, 发现MSJ1所在BAC上含有一个参与蓝光信号途径的基因AT5G64330, 其所编码的蛋白NPH3能够和蓝光受体PHOT1相互作用(表4), 推测可能是基因AT5G64330发生了突变, 随后对它进行测序。
| 表4 MSJ1基因功能预测 Table 4 Description of gene for MSJ1 |
对突变体phot1p2sa2中的AT5G64330基因进行测序。通过序列比对, 发现AT5G64330基因的第1252 bp发生了点突变, 由原来G变为A (图2-A)。即基因组编码区的第4个外显子上的1020碱基发生突变(图2-B)。蛋白质序列分析发现, 是三联子密码TGG突变为TGA (即由第255个氨基酸色氨酸突变为终止密码子)(图2-C), 即第255个氨基酸以后的编码丢失, 导致其功能发生巨大的差异, 拟南芥缺失强蓝光诱导的下胚轴向光弯曲反应。
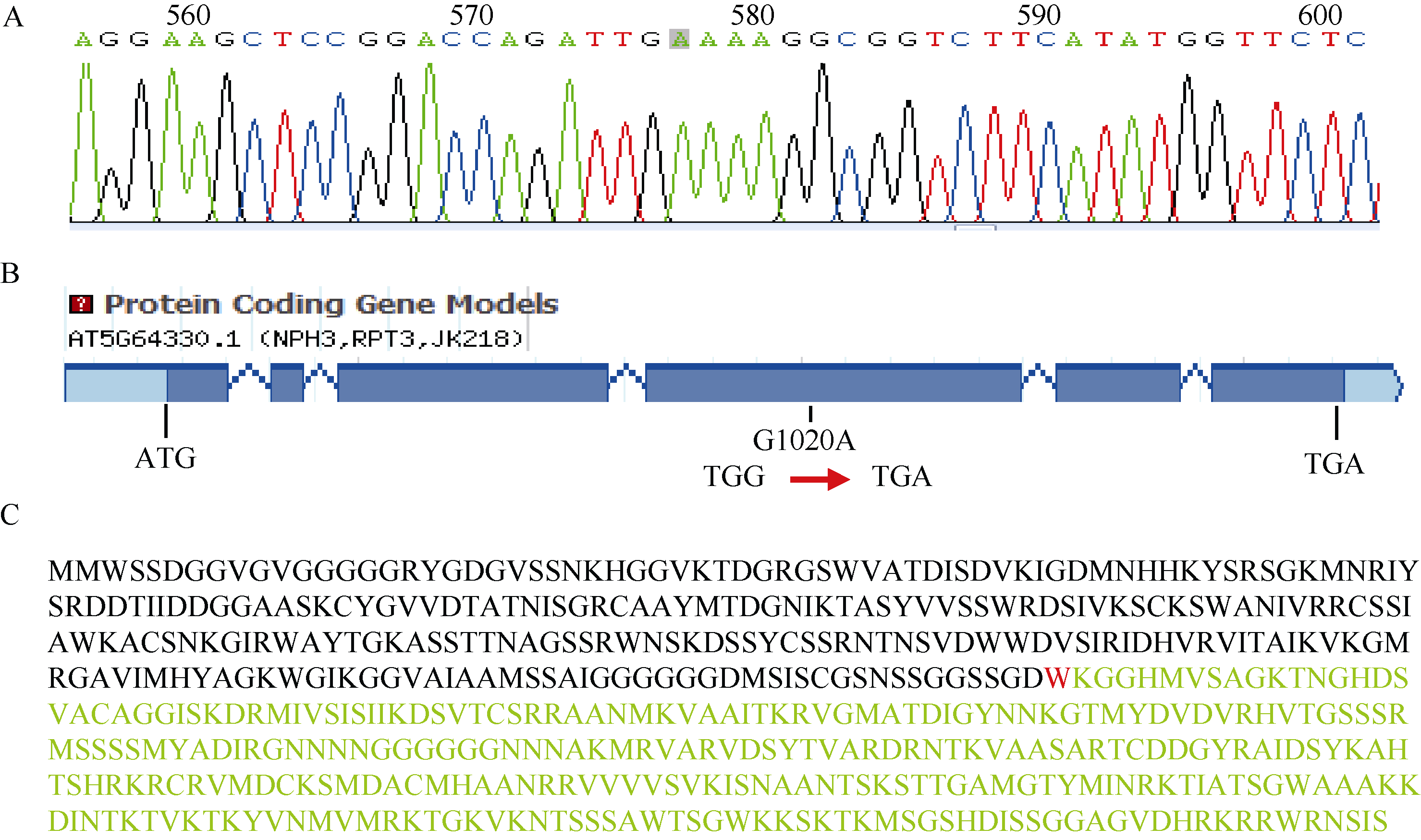 | 图2 P2SA2的定位及测序突变位点分析 A: P2SA2的突变位点; B: p2sa2突变体的突变分析; C: P2SA2基因编码蛋白序列及突变分析, 编码第255位色氨酸(红色W)密码子突变为终止密码子。Fig. 2 Map-based cloning of P2SA2gene and sequence of the mutant gene A: mutation of P2SA2 gene; B: mutation analysis of p2sa2; C: P2SA2 protein sequence. The 255th codon which encodes tryptophan (red W) was mutated to the terminator. |
以phot1、nph3-6突变体、phot1phot2双突变体以及phot1p2sa2突变体为材料, 比较下胚轴向光弯曲表型。如图3所示, 100 µ mol m-2 s-1蓝光单侧照射12 h, 突变体nph3-6表型类似于phot1p2sa2和phot1phot2突变体, 表现为强蓝光诱导的拟南芥下胚轴向光弯曲缺失, 而phot1突变体向光弯曲正常, 暗示突变体phot1p2sa2缺失强蓝光诱导的下胚轴向光弯曲, 可能是基因AT5G64330突变所致。
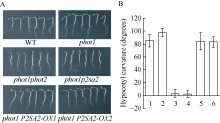
为进一步证明phot1p2sa2突变体表型确实是AT5G64330基因突变所致, 在phot1p2sa2突变体背景下, 构建了P2SA2的超表达植株。如图4所示, phot1p2sa2突变体背景下超表达P2SA2/NPH3 (AT5G64330)基因, 确实恢复了phot1p2sa2突变体下胚轴向光弯曲的生长, 表型类似于野生型(Col)和phot1突变体, 不同于突变体phot1phot2、phot1p2sa2缺失强蓝光诱导的拟南芥下胚轴向光弯曲(图4-A和图4-B)。上述结果进一步证实, phot1p2sa2突变体生理表型, 的确是P2SA2/NPH3 (AT5G64330)基因突变所致。
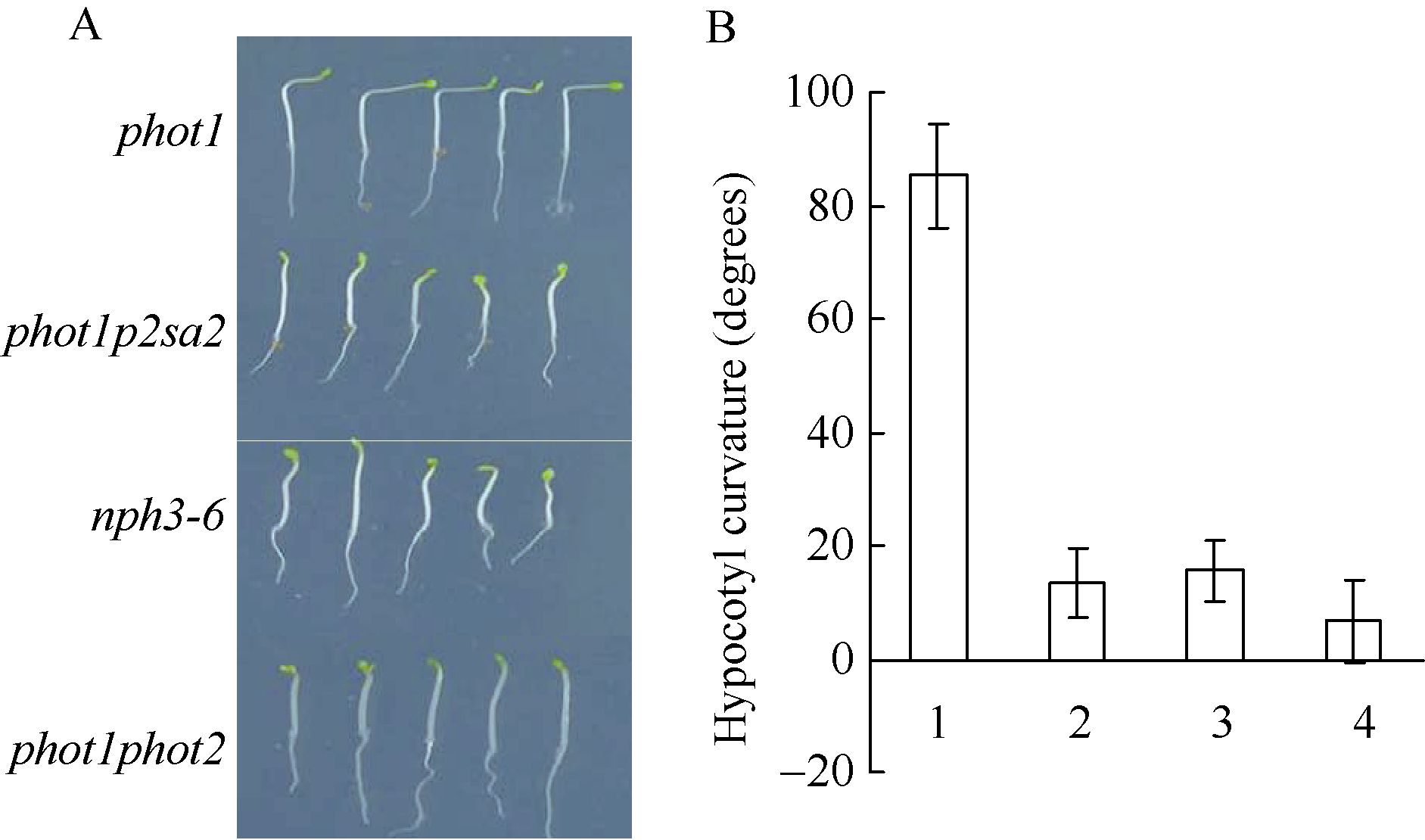 | 图3 NPH3调节强蓝光诱导的下胚轴弯曲度生长 A: 100 µ mol m-2 s-1蓝光单侧处理12 h后不同基因型拟南芥下胚轴向光弯曲表型; B: 不同基因型拟南芥下胚轴向光弯曲度统计。图中每个数据分别来自3次独立重复试验, 大约19~21颗苗的平均值± 标准误。其中1: phot1; 2: phot1p2sa2; 3:nph3-6; 4: phot1phot2。Fig. 3 Development of phototropic curvature in etiolated seedlings of indicated lines A: development of phototropic curvature in etiolated indicated lines at 100 µ mol m-2 s-1 blue light for 12 hours; B: hypocotyl curvatures were measured in indicated lines. 1:phot1; 2:phot1p2sa2; 3:nph3-6; 4:phot1phot2. Values are the means ± SD (n = 19-21). |
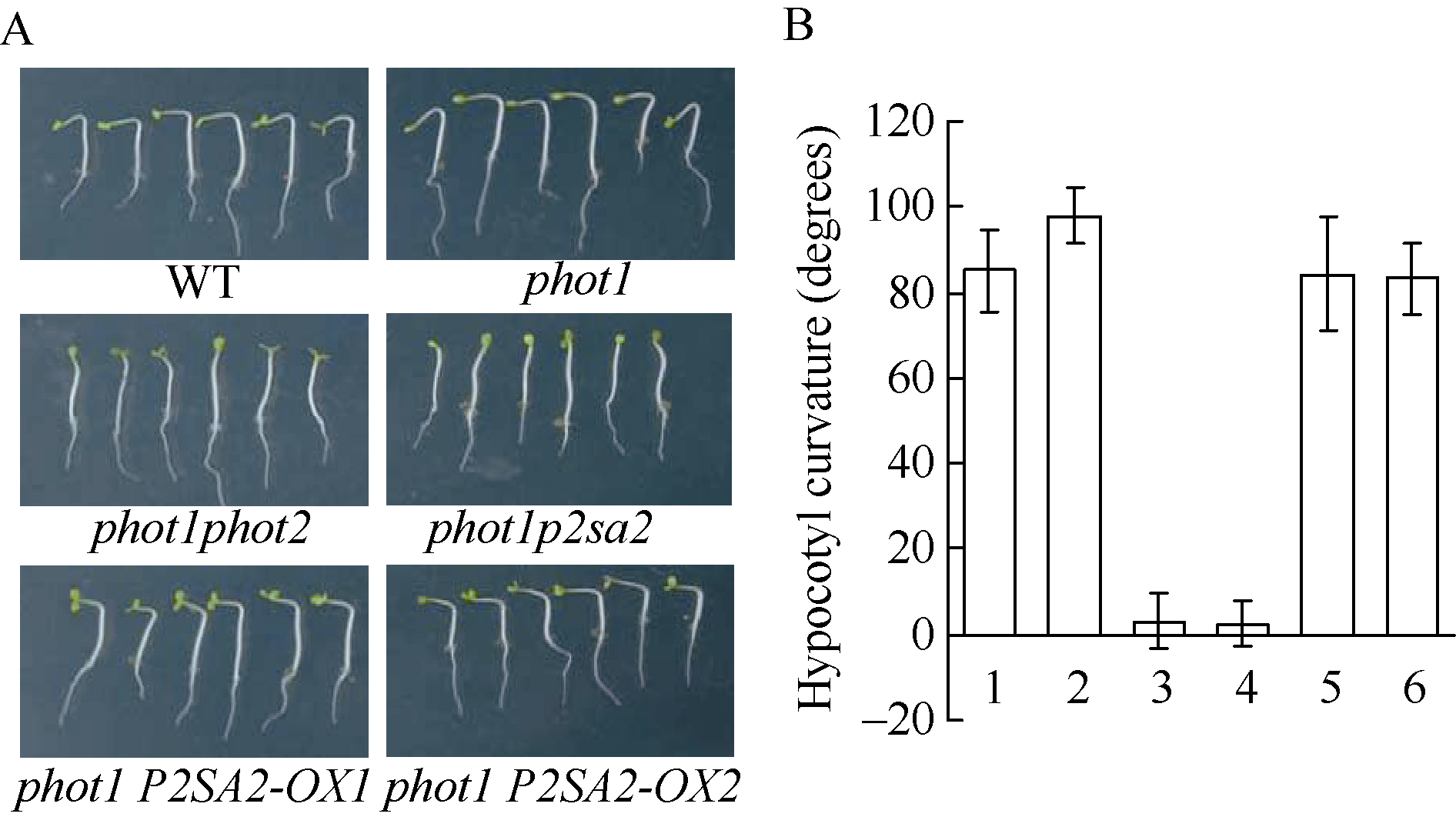 | 图4 P2SA2调节强蓝光诱导胚轴向光弯曲生长 A: 100 µ mol m-2 s-1蓝光单侧处理12 h不同基因型拟南芥下胚轴向光弯曲表型; B: 不同基因型拟南芥下胚轴向光弯曲度统计。图中每个数据分别来自3次独立重复试验, 大约18~21颗苗的平均值± 标准误。其中1: WT; 2: phot1; 3: phot1phot2; 4: phot1p2sa2; 5:phot1 P2SA2-OX1; 6:phot1 P2SA2-OX2。Fig. 4 Hypocotyl phototropism of p2sa2 mutant in etiolated seedlings was recovered by P2SA2 A: typical seedling phototropic curvature in indicated lines at 100 µ mol m-2 s-1 blue light for 12 hours; B: hypocotyl curvatures were measured in indicated lines. 1: WT; 2:phot1; 3:phot1phot2; 4:phot1p2sa2; 5:phot1 P2SA2-OX1; 6:phot1 P2SA2-OX2. Values are the means ± SD (n = 18-21). |
PHOT1和PHOT2既能通过不同的信号通路分别调节不同的生理反应, 又能以共同的信号分子调节下胚轴向光弯曲从而实现功能互补[21]。这种功能互补性限制了PHOT2调节下胚轴弯曲机制的研究。为此, 以拟南芥phot1为材料, 通过EMS诱变筛选PHOT2下游调节因子, 成功克隆到基因P2SA2。由于是以phot1突变体为背景材料, 因此相应突变体应该为phot1p2sa2。对phot1p2sa2突变体的遗传分析表明, phot1p2sa2是单基因隐性突变, 即该突变体的表型由P2SA2基因突变所致, 并不受PHOT1基因突变的干扰。为确定具体的突变位点, 首先对phot1p2sa2突变体进行图位克隆, 经过粗定位、细定位、猜基因、基因功能预测、测序等过程后, 最终认为P2SA2基因是定位于MSJ1这个BAC上的AT5G64330基因, 该基因1020 bp的G变为A, 三联子密码TGG突变为TGA, 最终导致基因功能缺失。该基因与已经报道的NPH3基因是等位基因。P2SA2基因突变后, 导致拟南芥缺失强蓝光诱导的下胚轴向光弯曲反应, 类似于phot1phot2双突变表型。由Christian Fankhauser (瑞士日内瓦大学)惠赠的P2SA2的T-DNA插入突变体nph3-6, 缺失强蓝光诱导的拟南芥下胚轴向光弯曲。结合我们获得的phot1p2sa2突变体表型回补的超表达阳性植株, 恢复拟南芥phot1p2sa2突变体下胚轴向光弯曲的表型, 证实该突变体缺失强蓝光诱导的拟南芥下胚轴向光弯曲的表型, 确实是NPH3(At5G64330)突变所致。
NPH3基因编码蛋白含有两个蛋白-蛋白互作结构域, 一个是BTB (broad complex, tramtrack, and bric-a-brac)/POZ (pox virus and zinc finger), 另一个是位于N和C末端的螺旋-螺旋结构域[11, 22], 可能是作为接头蛋白, 介导蛋白互作。暗处生长的拟南芥黄化幼苗, 其表达的NPH3蛋白被磷酸化, 弱蓝光照射可诱导其发生PHOT1依赖的去磷酸化反应[22], 调控生长素输出载体PIN1和输入载体AUX1[14, 15]的活性及定位, 导致下胚轴向光和背光侧中生长素的不对称分布, 诱导NPH4/ARF7介导的向光弯曲[23]。鉴于nph3突变体缺失任何强度光(弱光/强光)照下向光弯曲反应, 暗示NPH3蛋白可能是PHOT1和PHOT2介导向光性信号转导途径中的重要调节因子[24]。已有研究证明NPH3蛋白定位于细胞质膜, 体外体内均可与PHOT1和PHOT2相互作用[11, 13, 25]。鉴于强蓝光下phot1突变体下胚轴表现向光弯曲, 此时NPH3并未发生去磷酸化[26], 暗示NPH3的去磷酸化对于PHOT2介导的向光性并非必要。强蓝光下PHOT2如何调控NPH3活性介导下胚轴的向光弯曲反应机制并不清楚。近年来, 通过酵母三杂交, 找到了NPH3的另一个结合蛋白EHB1 (enhanced bending 1) 受到向光性和向重力刺激, ehb1突变体下胚轴弯曲度增强表明该蛋白是向光性和向重力性反应的负调控因子[27], 同时EHB1有可能是向光素或者生长素驱动Ca2+的内流进入囊泡运输途径的信号传导子[28]。NPH3有可能是通过与EHB1的互作调控生长素的不对称分布或是Ca2+的内流来调节PHOT2介导的强蓝光诱导的下胚轴向光弯曲反应。
通过图位克隆技术, 成功克隆基因P2SA2, 该基因的突变导致拟南芥缺失强蓝光诱导的下胚轴向光弯曲。测序比对和功能回补验证该突变体缺失强蓝光诱导的拟南芥下胚轴向光弯曲系AT5G64330基因的突变所致。该研究方法的建立将为强蓝光下PHOT2下游调节基因的筛选提供研究思路。蛋白P2SA2介导强蓝光诱导的拟南芥下胚轴向光弯曲的功能解析, 将为揭开PHOT2调节强蓝光诱导的下胚轴弯曲的机制提供重要理论基础。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|