


{kind=link}
{kind=link}
{kind=link}
{kind=link}
酵母双杂交检测
[高启国1, *  , 刘豫东
, 刘豫东1, * , 蒲全明1 , 张林成1 , 朱利泉2 , 王小佳1 ]
, 刘豫东|
|


* 第一作者联系方式: E-mail: gaoqg2004031@163.com, Tel: 023-68251974 **同等贡献(Contributed equally to this work)
Exo70A1是芸薹属作物柱头接受亲和花粉的必需因子, 可能参与了干性柱头水分分泌和花粉水合过程。本文从自交不亲和甘蓝A1柱头cDNA中获取了 BoExo70A1、 BoSEC3、 BoSEC10、 BoSEC15和 BoExo84基因的编码序列, 序列分析表明, BoExo70A1、 BoSEC3、 BoSEC10、 BoSEC15和 BoExo84基因分别与 AtExo70A1、 AtSEC3A、 AtSEC10、 AtSEC15B和 AtExo84B基因的cDNA序列高度同源, 其编码的5个蛋白均没有信号肽, BoSEC3蛋白含有一个典型的EF-hand钙离子结合结构域。将BoExo84蛋白拆分为2个片段(BoExo84-N和BoExo84-C), 并与BoSEC3、BoSEC10和BoSEC15蛋白编码序列一起分别亚克隆至pGADT7质粒, BoExo70A1蛋白编码序列亚克隆至pGBKT7质粒, 酵母双杂交检测结果表明, BoExo70A1蛋白能与BoSEC3、BoExo84-N蛋白互作, 但不能与BoSEC10、BoSEC15蛋白互作, 这为深入探讨BoExo70A1蛋白乃至整个胞泌复合体在甘蓝干性柱头接受自花花粉排斥异花花粉过程中的作用机制提供了依据。
Exo70A1 is necessary for Brassica stigma accepting the compatible pollen, and may play an important role in regulating the movement of water from the dry stigma to the pollen grain and compatible pollen hydration. Here, the coding sequences of BoExo70A1, BoSEC3, BoSEC10, BoSEC15, and BoExo84 genes were amplified from the stigma cDNA of the highly self-incompatible Brassica oleracea L. line A1. Sequence analysis showed that the cDNA sequences of BoExo70A1, BoSEC3, BoSEC10, BoSEC15, and BoExo84 genes were highly homologous to those of AtExo70A1, AtSEC3A, AtSEC10, AtSEC15B, and AtExo84B genes respectively. No signal peptide was found in their deduced protein sequences. BoSEC3 protein contained an EF-hand calcium-binding domain. The BoExo84 protein was divided into two fragments, BoExo84-N and BoExo84-C, and then their encoding sequences were subcloned into pGADT7 vector together with those of BoSEC3, BoSEC10, and BoSEC15 proteins respectively, whereas the encoding sequence of BoExo70A1 protein was subcloned into pGBKT7 vector for analysis of the proteins interaction by yeast two-hybid system. The results showed that BoExo70A1 protein interacted with BoSEC3 or BoExo84-N protein, but not with BoSEC10 or BoSEC15 protein.
甘蓝等芸薹属作物具有特异的干性柱头, 能识别自花或异花花粉, 只有当异花或不同单倍型的花粉落到柱头上后, 才能使柱头释放出水分和一些小分子物质, 完成授粉授精过程; 而自花或相同单倍型花粉落到柱头上后, 花粉与柱头蛋白互作后引发了柱头内自交不亲和信号传导过程, 最终导致花粉萌发受阻[1, 2, 3]。至今人们已对自花花粉与柱头互作早期事件分子机制研究较为清楚[3, 4, 5]。当自花花粉落到柱头上后, 花粉中S-位点半胱氨酸富集蛋白(S-locus cysteine-rich protein, SCR)与柱头S-位点受体激酶(S-locus receptor kinase, SRK)胞外域结合[6], 引起SRK构像的变化, 使其释放原本结合在SRK激酶结构域的类硫氧还蛋白1/2 (thioredoxin-h-like 1/2, THL1/2)[7]激发SRK激酶活性, 之后SRK与臂重复蛋白1 (arm repeat containing, ARC1)作用并使其磷酸化[8], 进而经过一系列目前尚未知晓的途径继续传递来自于细胞外信号, 最终导致花粉萌发受阻。
Samuel等[9]研究表明ARC1与Exo70A1作用, Exo70A1是柱头接受亲和花粉的必需因子, 过量表达Exo70A1在一定程度上降低材料的自交不亲和性。进一步研究表明Exo70A1参与亲和授粉后拟南芥和油菜柱头乳突细胞中可分泌型囊泡和多泡体向质膜花粉结合位点运输的调控[10], 高湿环境可部分恢复拟南芥AtExo70A1突变体柱头接受亲和花粉能力, 表明Exo70A1参与干性柱头水分分泌和花粉水合过程调控[11]。然而目前对于Exo70A1参与上述过程的分子机制尚未研究清楚。在真核生物中, 胞泌复合体是一个进化上高度保守的八聚体, Exo70A1是该复合体中一个亚基, 胞泌复合体可以将分泌型囊泡运输并束缚在细胞质膜的特定位点上[12, 13]。Há la等[14]研究表明拟南芥胞泌复合体亚基AtExo70A1与AtSCE3相互作用。本试验以高度自交不亲和甘蓝A1自交系开花前1~2 d的柱头cDNA为模板, 成功克隆了甘蓝胞泌复合体亚基BoExo70A1、BoSEC3、BoSEC10、BoSEC15和BoExo84的编码序列, 进行了生物信息学分析, 并利用酵母双杂交技术证明BoExo70A1可与BoSEC3和BoExo84-N相互作用, 以期为进一步探讨BoExo70A1在芸薹属作物柱头接受异花花粉和排斥自花花粉过程中作用的分子机制提供依据。
选取高度自交不亲和甘蓝A1高代自交系开花前1~2 d长势一致的花蕾, 取其柱头, 置-80℃冰箱备用, 用于提取柱头总RNA。
参照RNARep Pure植物总RNA试剂盒(TIANGEN, 北京)说明书提取柱头总RNA。依Primescript RT Reagent
Kit RR047A (TaKaRa, 大连)操作说明书反转录合成cDNA的第1条链, 置-20℃冰箱保存, 用于基因编码序列的克隆。
依Samuel等[9]公布的油菜BnExo70A1的cDNA序列(GenBank登录号为GQ503256), 在芸薹属基因组数据库(http://brassicadb.org/brad/index.php)进行BlastN检索, 获取其同源基因序列Bol010046, 同样利用拟南芥AtSEC3(AT1G47550)、AtSEC10(AT5G12370)、AtSEC15(AT4G 02350)和AtExo84 (AT5G49830)的CDS序列在芸薹属基因组数据库进行检索, 获取各自甘蓝同源基因的序列Bol044898、Bol043500、Bol006050和Bol035257, 然后依据Bol010046、Bol044898、Bol043500、Bol006050和Bol035257编码序列以及AtExo84的同源基因Bra037951序列, 利用Primer Primer 5软件设计引物(表1), 分别用于扩增甘蓝A1材料中胞泌复合体亚基BoExo70A1、BoSEC3、BoSEC10、BoSEC15和BoExo84的编码序列。
以反转录合成的柱头cDNA第1链为模板, 利用PrimeSTAR Max DNA聚合酶(TaKaRa, 大连)扩增各目的基因, 反应体系为25 μ L, 其中PrimerSTAR Max Premix 12.5 μ L、正反引物各1 μ L、cDNA 1 μ L、灭菌蒸馏水9.5 μ L。PCR循环参数为98℃ 2 min; 98℃ 10 s; 退火15 s; 72℃ 30 s; 35个循环; 72℃ 10 min (表1)。PCR产物经1.5%琼脂糖凝胶电泳后回收目的产物, 与pEASY-Blunt Simple克隆载体(TRANSGEN, 北京)连接后转入大肠杆菌DH5α , 将PCR鉴定的阳性克隆送华大基因公司测序。
利用DNAStar软件分析基因编码框及其推导编码氨基酸, 在美国国家生物技术信息中心(NCBI, http://blast. ncbi.nlm.nih.gov/Blast.cgi)、芸薹属基因组数据库(BRAD, http://brassicadb.org/brad/blastPage.php)和拟南芥信息资源数据库(TAIR, 网址: http://www.arabidopsis.org/wublast/ index2.jsp)利用BlastN在线软件分析基因序列相似性。用NCBI网站BlastP软件和PROSITE (http://prosite.expasy. org/)在线软件分析基因编码蛋白保守的结构域和功能位点, 利用SignalP (网址: http://www.cbs.dtu.dk/services/ SignalP/)在线软件分析蛋白信号肽序列, 通过Predict Protein (http://www.predictprotein.org/)在线软件预测蛋白亚细胞定位。
| 表1 基因扩增引物 Table 1 Primers for genes amplification |
依据获取的基因序列, 重新设计PCR引物并在各引物上引入相应的酶切位点(表2), 参照1.2中的PCR体系和循环参数扩增各目的基因片段, 其后对产物直接进行双酶切, 酶切产物后经1.5%琼脂糖凝胶电泳后回收目的片段, 与同样酶切的载体片段经Ligation high T4 DNA连接酶连接后转化大肠杆菌DH5α 。其中BoExo70A1亚克隆至酵母载体pGBKT7, BoSEC3、BoSEC10、BoSEC15亚克隆至酵母载体pGADT7。Há la等[14]将AtExo84亚克隆至载体pGBKT7, 转化酵母菌株后有极强的自激活活性, 为避免BoExo84自激活对蛋白质相互作用的影响, 本试验将BoExo84拆分为C端和N端2个片段, 分别亚克隆至载体pGADT7。获取的大肠杆菌DH5α 转化菌株, 经PCR鉴定后, 提取质粒进行双酶切鉴定, 将鉴定正确的菌株送华大公司测序。
| 表2 酵母重组质粒构建引物 Table 2 Primers for construction of yeast recombinant plasmid |
酵母宿主菌AH109感受态的制备和重组质粒pGBKT7- Exo70A1、pGADT7-SCE3、pGADT7-SEC10、pGADT7- SEC15、pGADT7-Exo84-N以及pGADT7-Exo84-C的毒性与自激活检测参照Matchmaker gal4 two-hybrid system3 (TaKaRa, 大连)操作说明书的步骤进行, pGBKT7-Exo70A1/ pGADT7-SEC3、pGBKT7-Exo70A1/pGADT7-SEC10、pGBKT7-Exo70A1/pGADT7-SEC15、pGBKT7-Exo70A1/ pGADT7-Exo84-N以及pGBKT7-Exo70A1/pGADT7-Exo84- C质粒组合转化感受态酵母AH109参照Matchmaker gal4 Two-hybrid System3操作说明书中小规模共转化的方法进行。相互作用的检测方法依据操作说明书调整如下, 将转化后的AH109菌株分别涂布在SD/-Leu/-Trp、SD/-Leu/-Trp/-His固体筛选培养基上30℃倒置培养; 挑取SD/-Leu/-Trp/-His培养基上的单菌落进行PCR鉴定(表2); 阳性克隆悬浮于0.5× YPDA (2 μ L)液体培养基中, 混匀后将悬浮液滴在SD/-Leu/-Trp/- His/-Ade/X-α -gal/25 mmol L-1 3-AT的固体筛选培养基上; 30℃倒置培养3~5 d, 观察菌落生长情况和颜色变化情况。
测序结果说明扩增的BoExo70A1编码序列为1917 bp、编码638个氨基酸, BoSEC3为2625 bp、编码875个氨基酸, BoSEC10为2430 bp、编码810个氨基酸, BoSEC15为2127 bp、编码709个氨基酸, BoExo84为2250 bp, 编码749个氨基酸。BlastN结果显示, BoExo70A1包含其从起始密码子ATG到终止密码子TAA的全部序列, 该基因与BnExo70A1 (GQ503256)、BoExo70A1 (Bra009523)、BrExo70A1 (Bra009523)及AtExo70A1 (AT5G03540.1)的相似性分别为99%、99%、97%和91%; 与BoSEC3 (Bol044898)比对, BoSEC3的3° 端少了33个碱基, BoSEC3与BrSEC3A (XM009120683)、BoSEC3A (Bol044898)和AtSEC3A (AR1G47550.1)相似性分别为99%、99%和93%; 与BoSEC10 (Bol043500)比对, BoSEC10在1845 bp处存在24个碱基的缺失, 其3° 端少了66个碱基, 与BraSEC10 (Bra008893)、BoSEC10 (Bol043500)和AtSEC10 (AT5G12370.1)相似性分别为97%、99%和91%。与BoSEC15 (Bol006050)比对, BoSEC15少了5° 端的210个碱基和3° 端的9个碱基, BoSEC15与BrSEC15B (Bra036274)、BoSEC15B (Bol006050)、AtSEC15B (AT4G02350.1)和AtSEC15A (AT3G56640.1)的相似应分别为95%、98%、84%和61%。BoExo84包含了其从起始密码子ATG到终止密码子TAA的全部序列, 该基因与BraExo84B(Bra037951)、BoSEC84B (Bol035257)和AtExo84B(AT5G49830.3)的相似性分别为98%、100%和92%。
BlastP结果表明BoExo70A1蛋白第271至第627位间氨基酸为Exo70家族保守结构域, BoSEC3蛋白第48至第144位间氨基酸为SEC3氮端PIP2结合PH结构域, 其第224至第868位间氨基酸是SEC3家族碳端保守结构域, BoSEC10蛋白第142至第810位间氨基酸是SEC10家族保守的结构域, BoSEC15蛋白第368至第645位间氨基酸为SEC15家族蛋白保守的结构域, BoExo84蛋白第27至第110位氨基酸为Vps51家族保守结构域。通过在线PROSITE软件分析BoSEC3的第553至第565位氨基酸为EF-hand钙离子结合结构域。经过SignalP 4.1在线软件分析表明BoExo70A1、BoSEC3、BoSEC10、BoSEC15和BoExo84蛋白均没有信号肽, 经Predict Protein在线软件预测, BoSEC3、BoSEC15和BoExo84被定位于细胞核, BoExo70A1和BoSEC10被定位于细胞质。
将BoExo70A1编码序列经NdeI/Sal I双酶切后亚克隆至pGBKT7质粒, BoSEC3、BoSEC10和BoSEC15编码序列分别经Sma I/ BamH I、Cla I/ BamH I和Nde I/ EcoR I双酶切后亚克隆至pGADT7质粒, 将BoExo84拆分成N端和C端连个部分, 其中N端包含了第2至第114位氨基酸, C端包含了第369至第749位氨基酸。重组酵母质粒双酶切结果(图1)表明均可以切出载体和目的基因片段, 且片段大小与理论值一致。重组酵母质粒测序结果显示各目的基因插入位点、插入方向以及编码框均正确, 说明可以进行后续相互作用验证。
 | 图1 酵母重组质粒双酶切电泳图M: Trans2K plus II DNA marker; 1: pGBKT7-BoExo70A1质粒Nde I/Sal I双酶切; 2: pGADT7-SEC3质粒Sma I/BamH I双酶切; 3: pGADT7-BoSEC10质粒Cla I/BamH I双酶切; 4: pGADT7-BoSEC15质粒Nde I/ EcoR I双酶切; 5: pGADT7-BoExo84-N质粒EcoR I/Xhol I双酶切; 6: pGADT7-BoExo84-C质粒EcoR I/Xhol I双酶切。Fig. 1 Double digestion patterns of recombinant plamidsM: Trans2K plus II DNA marker; 1: plasmid pGBKT7-BoExo70A1 digested byNde I/Sal I; 2: plasmid pGADT7-SEC3 digested by Sma I/BamH I; 3: plasmid pGADT7-BoSEC10 digested by Cla I/BamH I; 4: plasmid pGADT7-BoSEC15 digested byNde I/ EcoR I; 5: plasmid pGADT7-BoExo84-N digested by EcoR I/Xho I; 6: plasmid pGADT7-BoExo84 digested by EcoR I/Xho I. |
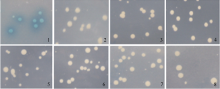
参照酵母双杂交试剂盒说明书将构建的酵母重组质粒pGBKT7-Exo70A1、pGADT7-SEC3、pGADT7-SEC10、pGADT7-SEC15、pGADT7-Exo84-N以及pGADT7-Exo84- C分别转化酵母AH109感受态, 其中pGBKT7-Exo70A1转化的菌株涂在SD/-Trp平板上, 其余5个重组质粒转化的菌株涂在SD/-Leu平板上, 30℃倒置培养3~5 d, 以pGBKT7质粒转化的AH109在SD/-Trp平板上的菌落作为对照, 分析重组蛋白对酵母菌落生长影响, 结果表明各重组质粒转化的酵母菌株在相应的平板上均可以长出菌落且大小基本一致, 表明表达的BoExo70A1、BoSEC3、BoSEC10、BoSEC15、BoExo84-N和BoExo84-N蛋白均没有毒性(图2)。参照酵母双杂交试剂盒说明书, 将pGBKT7-Exo70A1与pGADT7空载体、其余5个酵母重组质粒分别与pGBKT7空载体组合共转化酵母AH109感受态, 转化后的菌液涂在SD/-Trp/-Leu/X-α -Gal平板上, 30℃倒置培养3~5 d, 以酵母试剂盒附带的阳性和阴性对照质粒组合转化平板为参照, 观察菌落的显色情况, 结果(图3)显示只有阳性对照质粒组合转化后的菌株显蓝色, 其余的全部为白色菌落, 说明表达的BoExo70A1、BoSEC3、BoSEC10、BoSEC15、BoExo84-N和BoExo84-N蛋白均没有自激活活性。
将酵母重组质粒pGBKT7-Exo70A1分别与pGADT7- SEC3、pGADT7-SEC10、pGADT7-SEC15、pGADT7- Exo84-N以及pGADT7-Exo84-C两两组合后转化酵母AH109感受态, 涂在SD/-Leu/-Trp培养基上, 30℃倒置培养3~5 d, 提取菌落用通用引物进行PCR检测, 选取能扩出2个目的基因条带且直径大于2 mm的菌落, 通过改良的方法转接到SD/-Leu/-Trp/-His/-Ade/X-α -gal/25 mmol L-1 3-AT的固体筛选培养基上; 30℃倒置培养3~5 d, 观察菌落生长情况和颜色变化情况。结果表明阳性对照、pGBKT7-Exo70A1/pGADT7-SEC3、pGBKT7-Exo70A1/ pGADT7-Exo84-N组合转化的菌株在SD/-Leu/-Trp/-His/- Ade/X-α -gal/25 mmol L-1 3-AT平板上菌落能生长, 且菌落呈现蓝色(图4), 说明BoExo70A1与BoSEC3、BoExo70A1与BoExo84-N之间能相互作用; 而pGBKT7- Exo70A1/pGADT7-SEC10、pGBKT7-Exo70A1/pGADT7- SEC15以及pGBKT7-Exo70A1/pGADT7-Exo84-C的3对组合与阴性对照转化的酵母菌株一样在SD/-Leu/-Trp/- His/-Ade/X-α -gal/25 mmol L-1 3-AT平板上均不能生长, 且由于不能激活下游Ade基因的表达导致菌落呈现微微的红色(图4), 表明BoExo70A1与BoSEC10、BoSEC15和BoExo84-C蛋白均不能相互作用。
 | 图2 酵母重组质粒毒性检测1: pGBKT7酵母转化子在SD/-Trp平板; 2: pGBKT7-BoExo70A1酵母转化子在SD/-Trp平板; 3: pGADT7-BoSEC3酵母转化子在SD/-Leu平板; 4: pGADT7-BoSEC10酵母转化子在SD/-Leu平板; 5: pGADT7-BoSEC15酵母转化子在SD/-Leu平板上; 6: pGADT7-BoExo84-N酵母转化子在SD/-Leu平板; 7: pGADT7-BoExo84-C酵母转化子在SD/-Leu平板。Fig. 2 Test of toxicity of yeast recombinant plasmids1: pGBKT7 yeast transformants on SD/-Trp plate; 2: pGBKT7-BoExo70A1 on SD/-Trp plate; 3: pGBKT7-BoSEC3 on SD/-Leu plate; 4: pGBKT7-BoSEC10 on SD/-Leu plate; 5: pGBKT7-BoSEC15 on SD/-Leu plate; 6: pGBKT7-BoExo84-N on SD/-Leu plate; 7: pGBKT7-BoExo84-C on SD/-Leu plate. |
 | 图3 酵母重组质粒自激活检测Fig. 3 Test of autoactivation for yeast recombinant plasmids1: pGBKT7-p53× pGADT7-T, 2: pGBKT7-Lam× pGADT7-T; 3: pGBKT7-BoExo70A1× pGADT7; 4: pGBKT7× pGADT7-BoSEC3; 5: pGBKT7× pGADT7-BoSEC10; 6: pGBKT7× pGADT7-BoSEC15; 7: pGBKT7× pGADT7-BoExo84-N; 8: pGBKT7× pGADT7-BoExo84-C. |
 | 图4 酵母中Exo70A1与其余5个蛋白相互作用分析Fig. 4 Analysis of the interaction of BoExo70A1 with the other five proteins in yeast1: negative control; 2: positive control; 3: pGBKT7-BoExo70A1× pGADT7-BoSEC3; 4: pGBKT7-BoExo70A1× pGADT7-BoSEC10; 5: pGBKT7-BoExo70A1× pGADT7-BoExo84-C; 6: pGBKT7-BoExo70A1× pGADT7-BoSEC15; 7: pGBKT7-BoExo70A1× pGADT7-BoExo84-N. |
在自交不亲和信号传导分子机制的研究中, Samuel等[9]过量表达ARC1互作蛋白Exo70A1部分打破材料的自交不亲和性, 反义干扰Exo70A1表达会降低柱头接受亲和花粉的能力, 表明Exo70A1可能是自花和异花在柱头内引发的2种信号通路的交叉点。通过透视电镜观察结果显示Exo70A1参与了拟南芥和琴叶拟南芥柱头乳突细胞内可分泌型囊泡以及油菜柱头乳突细胞内多泡体向细胞质膜的运输和富集过程调控, 自交不亲和信号传导过程可能通过ARC1泛素化降解Exo70A1蛋白, 进而使不亲和授粉后柱头乳突细胞内可分泌型囊泡或者多泡体分布在细胞质, 而不能向花粉结合位点的细胞质膜运输和富集[10]。高湿环境能部分恢复拟南芥Exo70A1突变体接受亲和花粉能力的研究结果, 进一步表明该蛋白参与柱头乳突细胞水分分泌和花粉水合过程的调控[11]。Iwano等[15]研究表明异花授粉将会引发自交不亲和芜菁柱头乳突细胞内肌动蛋白聚合, 而自花授粉将导致肌动蛋白的重组和解聚, 肌动蛋白解聚药品细胞松弛素D处理后将会抑制亲和花粉的水合和萌发, 同时自花授粉将扰乱柱头乳突细胞内液泡分布, 而异花授粉后将引起液泡向花粉结合位点重新排列, 表明肌动蛋白的聚合与解聚可能直接影响了柱头乳突细胞内囊泡的运输。本试验获取了甘蓝胞泌复合体BoExo70A1、BoSEC3、BoSEC10、BoSEC15和BoExo84亚基编码序列, 并利用酵母双杂交技术证明BoExo70A1与BoSEC3、BoExo84-N相互作用。
在出芽酵母中, SEC3可以作为细胞质膜上极性分泌位点的标志[16], SEC3和Exo70两个亚基通过非肌动蛋白束依赖的方式定位在细胞质膜上, 其余的胞泌复合体亚基与可分泌型囊泡束缚在一起, 并在肌球蛋白V型蛋白Myo2p作用下沿着肌动蛋白束移动, 一旦可分泌型囊泡被移动到肌动蛋白束末端, 与囊泡束缚在一起的胞泌复合体亚基与定位在细胞质膜上的SEC3和Exo70亚基作用, 进而将可分泌型囊泡束缚在细胞质膜上[17, 18]。在裂殖酵母中, SEC3和Exo70均可以将其他胞泌复合体亚基定位在细胞质膜上, 且两者具有功能冗余性[19]。但是与出芽酵母中SEC3和Exo70亚基独立于胞泌复合体, 提前定位在细胞质膜上不同的是, 裂殖酵母中包括SEC3和Exo70在内的所有胞泌复体亚基与可分泌型囊泡束缚在一起, 在肌球蛋白V型蛋白作用沿着肌动蛋白束移动, 进而将可分泌型囊泡定位在细胞质膜特定位点上[19]。上述两种类型的酵母研究结果表明, SEC3和Exo70亚基在可分泌型囊泡束缚在细胞质膜特定位点过程中起了关键作用, 且两种酵母中胞泌复合体运输囊泡的过程都依赖于肌动蛋白束。此外, 出芽酵母中Exo84蛋白对于胞泌复合体的聚合以及极性定位均起了关键作用[20]。
参照上述酵母中胞泌复合体运输可分泌型囊泡调控机制的研究结果, 下一步研究的重点是利用透视电镜[11]、罗丹明鬼笔环肽染色[15]和荧光蛋白标记[21]的方法分别观察自交不亲和甘蓝自花和异花授粉后柱头乳突细胞内可分泌型多泡体、肌动蛋白和胞泌复合体各亚基分布的动态变化, 以便深入解析BoExo70A1乃至整个胞泌复合体各亚基在柱头响应自花和异花授粉过程中的作用机制。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|