


{kind=link}
{kind=link}
{kind=link}
{kind=link}
三种禾谷类作物强、弱势粒淀粉粒形态与粒度分布的比较
[徐云姬, 李银银, 钱希旸, 王志琴, 杨建昌*  ]
]
]
|
|


第一作者联系方式: E-mail: xuyunji19881004@163.com
本研究以水稻品种武运粳24和扬两优6号、小麦品种扬麦16和宁麦13及玉米品种登海11和农乐988为试验材料, 分别提取其成熟期强、弱势粒的淀粉粒, 观察比较不同作物及其强、弱势粒间淀粉粒形态和淀粉粒数量、体积和表面积分布特性。结果表明, 3种禾谷类作物间淀粉粒形态大小差异明显, 粒径表现为玉米>小麦>水稻。水稻淀粉粒呈有棱角的无规则状, 小麦淀粉粒呈透镜体状或球体状, 玉米淀粉粒呈椭球体状、多面体状或圆球体状。水稻和玉米淀粉粒数量、体积和表面积分别成单峰、三峰和双峰分布; 小麦淀粉粒数量呈单峰分布, 体积呈微弱的四峰分布, 表面积呈三峰分布。水稻、小麦和玉米淀粉粒按各自粒径不同人为划分为小淀粉粒、中淀粉粒和大淀粉粒, 分界线分别为1.5 μm和20.0 μm、5.0 μm和50.0 μm、4.0 μm和50.0 μm。3种作物籽粒淀粉粒的总体积主要决定于中淀粉粒体积。3种作物的强、弱势粒间小淀粉粒粒度分布比例及中淀粉粒所占数量比例没有明显差异, 但各作物强势粒的中淀粉粒所占的体积和表面积比例均显著高于弱势粒, 大淀粉粒的分布比例低于弱势粒。强、弱势粒的中淀粉粒所占体积比例与其淀粉积累量和粒重的高低变化趋势一致。表明淀粉粒体积是决定粒重的一个重要因素, 增加弱势粒的中淀粉粒体积或减小大淀粉粒体积可望增加其粒重。
Using two rice cultivars, two wheat cultivars, and two maize cultivars, we extracted starch granules to observe the morphological characteristics and to compare the differences in the starch granule number, volume and surface area distributions among the three crops and between superior and inferior grains of each cultivar. The results showed that significant differences were observed in starch granule morphology and size among the three crops. The diameter of starch granule showed an order of maize > wheat > rice. Starch granules extracted in rice showed an irregular shape, the starch granules in wheat were lenticular-shaped and spherical-shaped. The starch granules in maize grain were mainly polyhedral or irregular and spherical in shape. Distributions of starch granule number, volume and surface area were changed in a typical unimodal-peak curve, a triple-peak curve and a typical bimodal-peak curve, respectively, in grains of rice and maize. Distributions of starch granule number, volume and surface area in wheat grains displayed a typical unimodal-peak curve, a four-peak curve and a triple-peak curve, respectively. According to starch granule diameters, all the starch granules were classified into small, medium, and large granules in this study. The thresholds for separating the starch granules were 1.5 μm and 20.0 μm, 5.0 μm and 50.0 μm, 4.0 μm and 50.0 μm, respectively, in rice, wheat and maize. The total volume of starch granules in grains of the three cereal crops was mainly determined by the volume of medium starch granules. No significant difference was observed in the proportions of small starch granule size and medium starch granule number between superior and inferior grains of the three cereal crops. But the volume and surface area of medium starch granules were larger in superior grains than in inferior ones. The percentage of large starch granules was greater in the inferior than in the superior. Changes in the volume of medium starch granules in both superior and inferior grains were consistent with those in starch accumulation and grain weight. The results suggest that the starch granule volume is an important factor determining grain weight. Increasing the volume of medium starch granules or reducing the volume of large starch granules would increase the weight of inferior grains.
籽粒重量是作物产量的重要组成部分, 籽粒充实的程度直接关系到粒重和产量。水稻、小麦和玉米等禾谷类作物的粒重又会因其在穗上着生的位置不同而有较大差异, 通常分为强势粒和弱势粒两种类型[1, 2, 3]。一般而言, 水稻的强势粒位于稻穗上部一次枝梗上[1], 小麦的强势粒位于麦穗中部[2], 玉米的强势粒位于果穗中下部[3]。强势粒灌浆快、充实好、粒重高。水稻的弱势粒位于稻穗下部二次枝梗上[1], 小麦的弱势粒位于麦穗的基部和上部或一个小穗的中部[2], 玉米的弱势粒位于果穗上部[3]。弱势粒灌浆慢、充实差、粒重低。禾谷类作物籽粒成分主要是淀粉, 籽粒灌浆过程实质上是淀粉合成的过程。因此, 强、弱势籽粒间的粒重差异主要归因于淀粉生物合成和累积量的不同[4, 5, 6]。据Jiang等[4]报道, 小麦强势粒之所以较弱势粒具有较高粒重, 是因为其较高的淀粉积累速率和较强的蔗糖-淀粉代谢途径关键酶活性。他们的研究表明, 成熟期小麦强势粒中直链淀粉和支链淀粉含量显著高于弱势粒[4]。玉米中也有类似报道, 玉米果穗上部籽粒(弱势粒)中的淀粉含量明显低于中下部籽粒(强势粒)[6]。
籽粒中淀粉是以半晶体状态的淀粉粒形式储存于胚乳中。淀粉粒的粒径从1 μ m到100 μ m不等, 形状各异(如球体状、透镜状、多面体和不规则状等), 大小分布特征也有差异(如单峰分布、双峰分布或三峰分布) [7, 8]。前人指出, 小麦成熟期籽粒胚乳中存在2种类型淀粉粒, 规定为A型大淀粉粒和B型小淀粉粒, 二者的分界线为9.9 μ m [9, 10]。Peng等[11]也观察到, 成熟期小麦胚乳中含有A型淀粉粒(10~35 μ m)和B型淀粉粒(1~10 μ m)。后来, 有研究者又发现了C型淀粉粒, 它们较B型淀粉粒形成晚, 并且粒径更小 [12]。据报道, 玉米淀粉粒被分为3种类型, 即小淀粉粒组(< 2 μ m)、中淀粉粒组(2~15 μ m)和大淀粉粒组(> 15 μ m)[13]。禾谷类作物淀粉粒度的数量分布、体积分布和表面积分布等, 不仅受遗传因素控制[13, 14, 15, 16], 而且受环境条件的影响[17, 18, 19]。但已有的研究多集中于单种作物或单种作物不同穗位籽粒中淀粉粒的研究, 缺乏对不同作物间及同一作物穗上不同部位间籽粒淀粉粒形态及粒度分布特征的比较分析。本研究观察比较3种典型禾谷类作物的强、弱势籽粒中淀粉粒形态及粒度分布特征, 以期从淀粉粒的层面上进一步阐明禾谷类作物强、弱势籽粒间淀粉积累及粒重差异的原因。
水稻试验于2012年和2014年在扬州大学实验农场进行。供试品种为武运粳24和扬两优6号, 其直链淀粉含量分别为14.6%和16.5%。试验地前茬均为小麦, 土壤为沙壤土, 耕作层含有机质2.02%、有效氮103.2 mg kg-1、速效磷24.5 mg kg-1、速效钾85.6 mg kg-1。5月10日至11日播种, 6月9日至10日移栽。株、行距为15 cm× 25 cm, 武运粳24双本栽, 扬两优6号单本栽, 小区面积为12.2 m × 5.6 m, 随机区组排列, 重复3次。全生育期施尿素折合纯氮240 kg hm-2, 按基肥∶ 分蘖肥∶ 穗肥=5∶ 2∶ 3施用。基施过磷酸钙(含P2O5 13.5%) 445 kg hm-2和氯化钾(含K2O 62.5%) 150 kg hm-2。其余田间管理同当地常规高产栽培。
小麦试验于2013年10月— 2014年5月在扬州大学江苏省作物遗传生理重点实验室试验农场进行。供试品种为普通小麦扬麦16和宁麦13, 其粗蛋白含量分别为14.2%和12.5%, 湿面筋含量分别为32.2%和27.6%。试验地前茬作物为水稻, 土壤质地为沙壤土, 含有机质2.02%, 有效氮为105.0 mg kg-1, 速效磷34.2 mg kg-1, 速效钾68.0 mg kg-1。2013年10月28日人工条播, 基本苗225万 hm-2, 行距30 cm, 三叶期定苗, 照高产栽培进行肥料运筹, 按基肥∶ 分蘖肥∶ 拔节肥∶ 孕穗肥=4∶ 2∶ 2∶ 2施用。全生育期施用尿素折合纯氮180 kg hm-2, 播种前一次性施过磷酸钙(含P2O5 13.5%)折合纯磷90 kg hm-2和氯化钾(含K2O 52%)折合纯钾90 kg hm-2, 其余管理措施同大田高产栽培。小区面积为12.2 m × 5.6 m, 随机区组排列, 重复3次。
玉米试验于2012年和2014年在扬州大学实验农场大田进行。供试品种为普通玉米杂交种登海11和农乐988, 其粗淀粉含量分别为71.6%和73.2%。试验地土壤类型为沙壤土, 土壤含有机质22.7 g kg-1、速效氮96.5 mg kg-1、速效磷20.4 mg kg-1、速效钾120.0 mg kg-1。2012年的试验于3月25日播种, 7月19日收获。2014年试验4月1日播种, 7月23日收获。两年玉米的株、行距均为37 cm × 55 cm, 三叶期间苗定苗至4.91万株 hm -2。播种前施用尿素、过磷酸钙和氯化钾, 分别折合纯氮42 kg hm-2、P2O5 76 kg hm-2 和K2O 95 kg hm-2。在大喇叭口期和吐丝期分别追施尿素折合纯氮27 kg hm-2和63 kg hm-2。试验小区面积均为235 m2, 重复2 次。其他管理措施同当地玉米高产栽培。
于水稻抽穗期选择大小基本一致的穗子150个挂牌, 标记部分穗颖花开花日期。成熟期取120个挂牌稻穗(每小区40个)用于剥取强、弱势粒。强势粒为穗上第1、第2天开花的颖花, 着生在穗顶部一次枝梗, 弱势粒为穗上最后2 d开花的颖花, 着生在穗基部二次枝梗。
于小麦开花期选择大小基本一致的穗子150个挂牌, 标记部分穗颖花开花日期。成熟期取120个挂牌麦穗(每小区40个)用于剥取强、弱势粒。强势粒为麦穗中部小穗(从基部往上数起的第4~第12小穗)的第1、第2位籽粒, 弱势粒为穗上部和穗下部小穗的全部籽粒。
玉米吐丝期选择同日吐丝且健壮一致的玉米10株挂牌标记。成熟期取挂牌果穗6个(每小区3个), 将果穗分为上、中、下三部分, 去除各分界处的边际籽粒后, 将中、下部籽粒混合作为强势粒, 取玉米果穗上部籽粒作为弱势粒。
1.2.1 强、弱势籽粒重测定
取强、弱势籽粒各400粒烘干至恒重, 称重后计算单粒重。将称完重的籽粒分别磨成粉末用于淀粉含量的测定和淀粉粒的提取。水稻籽粒烘干去除颖壳后再磨成粉。
1.2.2 强、弱势籽粒中淀粉含量的测定
上述烘干的强、弱势籽粒粉末过100目筛, 采用双波长比色法[20]测定直链淀粉和支链淀粉含量, 淀粉含量为直链淀粉和支链淀粉含量之和。
1.2.3 淀粉粒提取与纯化
参照Peng等[21]和Ji等[22]的方法并作修改提取淀粉粒。分别取成熟期强、弱势籽粒粉末5~10 g放入50 mL离心管中, 按50 mg g-1的比例各自加入适量碱性蛋白酶和25 mL NaOH溶液(pH 9~10), 42℃下振荡混匀24 h。然后将淀粉匀浆过200目筛, 滤液经4000× g离心20 min, 去上清液, 并刮掉淀粉表层黄色物质。用去离子水悬浮淀粉沉淀后4000× g离心20 min, 重复此步骤3~4次洗去离子与杂质。然后依次用95%的乙醇、氯仿和甲醇混合液(1∶ 1, v/v)、甲醇和丙酮混合液(1∶ 1, v/v)各洗涤2~3次去脂。将提取物质70℃烘至半干, 用玻璃棒搅匀后过200目筛得淀粉, 收集并于4℃冰箱密封保存备用。
1.2.4 淀粉粒形态观察
取2 mg提取的淀粉溶于5 mL无水乙醇中, 分别吸取1~2滴淀粉悬浊液加在样品台上, 25℃吹干后用离子溅射仪镀金膜, 于扫描电子显微镜(ESEM XL-30, Philips, the Netherlands)下观察与拍照。设置的电压和电流分别为20 kV和150 mA, 分别于5000× 、2500× 和2500× 放大倍数下观察水稻、小麦和玉米淀粉粒形态。
1.2.5 淀粉粒度分布测定
用英国马尔文公司的MS-2000激光粒度分析仪分析淀粉粒。取50 mg淀粉放入10 mL离心管, 加5 mL无水乙醇悬浮, 旋涡混匀后置4℃下1 h, 每10 min振荡1次, 在吸取样品前摇匀, 吸取2 mL左右转移至激光衍射粒度分析仪的装有无水乙醇的分散盒中, 测量淀粉粒数量、体积及表面积分布状况。采用仪器本身携带的软件获取结果, 以百分比表示。
采用Microsoft Excel 2003和SPSS16.0统计软件分析试验数据, 用SigmaPlot 10.0作图。用最小显著差法(LSD0.05)检验平均数。小麦为1年的试验数据。水稻和玉米因2年的结果趋势一致, 所以用2年的平均数。
由表1可知, 籽粒的重量表现为玉米> 小麦> 水稻。3种作物强势粒的最终淀粉积累量及其粒重均显著高于弱势粒。强、弱势粒间粒重的差异以玉米为最大, 其次为水稻, 小麦为最小(表1)。
| 表1 成熟期3种禾谷类作物强、弱势粒淀粉积累量及其粒重 Table 1 Grain weight and starch accumulation in superior and inferior grains of the three cereal crops at maturity |
由图1可见, 3种禾谷类作物间淀粉粒形态差异明显。水稻的淀粉粒明显小于小麦和玉米。水稻的淀粉粒形状不规则, 含有棱角(图1-A, B)。小麦的淀粉粒主要表现为透镜体状和圆球体状, 且表面光滑(图1-C, D)。玉米的淀粉粒主要成多面体状、椭球体状或球体状(图1-E, F)。
 | 图1 水稻(A, B: 放大倍数5000× )、小麦(C, D: 放大倍数2500× )和玉米(E, F: 放大倍数2500× )淀粉粒的形态特征Fig. 1 Scanning electron microscopy (SEM) showing the starch granule in rice (A, B: 5000× ), wheat (C, D: 2500× ) and maize (E, F: 2500× ) |
图2显示, 无论强势粒或弱势粒, 淀粉粒的数量分布均呈现单峰变化趋势, 且峰值对应的粒径在0.5 μ m左右, 体积分布表现为有一主峰的三峰分布, 其主峰峰值对应的粒径在6.0 μ m左右, 三峰分界对应的粒径分别在1.5 μ m左右和20.0 μ m左右。表面积分布表现为典型的双峰分布, 2个峰值对应的粒径分别在1.0 μ m左右和4.0 μ m左右。尽管强、弱势粒间的淀粉粒度分布规律极度相似, 但不同粒径范围内的数量、体积或表面积均存在或多或少的差异。两供试水稻品种的表现趋势一致( 图2)。
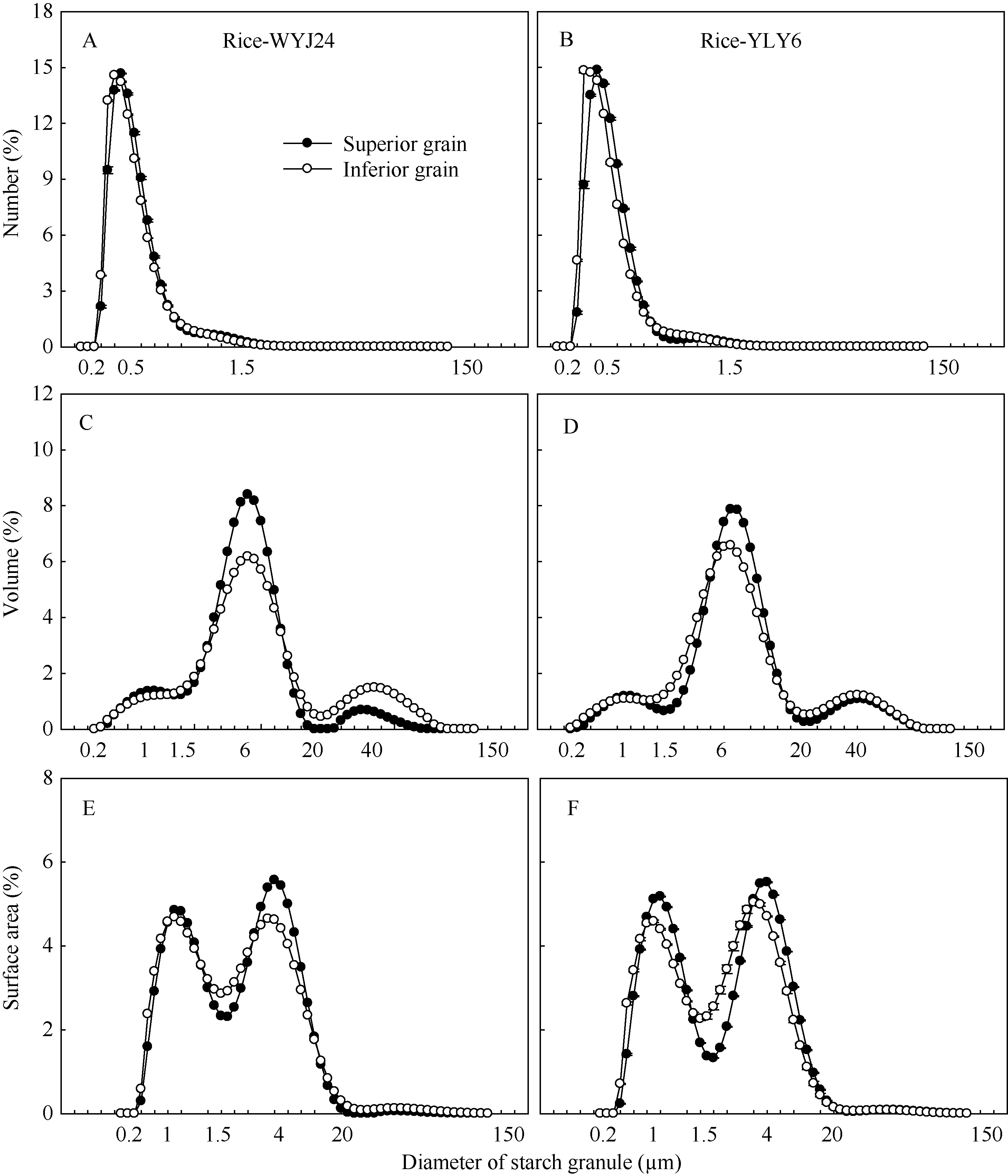 | 图2 两供试水稻品种强、弱势粒中淀粉粒的数量(A, B)、体积(C, D)和表面积(E, F)分布Fig. 2 Number (A, B), volume (C, D), and surface area (E, F) distributions of starch granule in the superior and inferior grains of two rice cultivars |
图3显示, 小麦强、弱势籽粒淀粉粒的数量呈单峰分布, 且峰值对应的粒径在0.5 μ m左右, 体积分布表现为微弱的四峰分布, 其主峰对应的峰值在20.0 μ m左右, 四峰分界对应的粒径分别在1.5 μ m左右、5.0 μ m和50.0 μ m左右; 表面积表现为三峰分布, 两侧主要峰值对应的粒径分别在0.5 μ m左右和15.0 μ m左右。尽管小麦强、弱势粒间的淀粉粒度分布规律也极度相似, 但不同粒径范围内的数量、体积或表面积均存在或多或少的差异。两供试小麦品种的趋势相同( 图3)。
 | 图3 两供试小麦品种强、弱势粒中淀粉粒的数量(A, B)、体积(C, D)和表面积(E, F)分布Fig. 3 Number (A, B), volume (C, D), and surface area (E, F) distributions of starch granule in the superior and inferior grains of two wheat cultivars |
图4显示, 无论强势粒或弱势粒, 玉米淀粉粒的数量分布呈单峰变化趋势, 且峰值对应的粒径在0.5 μ m左右, 体积分布表现为有一主峰的三峰分布, 主峰峰值对应的粒径在15.0 μ m左右, 三峰间的分界对应的粒径分别在4.0 μ m左右和50.0 μ m左右, 而表面积分布也表现为典型的双峰分布, 两个峰值对应的粒径分别在0.5 μ m左右和12.0 μ m左右。尽管强、弱势粒间的淀粉粒度分布规律也基本相同, 但不同粒径范围内的数量、体积或表面积均存在或多或少的差异。两供试玉米品种的表现趋势一致( 图4)。
 | 图4 两供试玉米品种强、弱势粒中淀粉粒的数量(A, B)、体积(C, D)和表面积(E, F)分布Fig. 4 Number (A, B), volume (C, D), and surface area (E, F) distributions of starch granule in the superior and inferior grains of two maize cultivars |
由图2至图4可以看出, 水稻、小麦和玉米籽粒中绝大部分的淀粉粒对应粒径大小分别在0.2~ 1.5 μ m、0.2~5.0 μ m和0.2~50.0 μ m之间。淀粉粒平均粒径表现为玉米> 小麦> 水稻, 这与它们的粒重大小相一致。
由表2得知, 两供试水稻品种的小淀粉粒(粒径< 1.5 μ m)和中淀粉粒(粒径为1.5~20 μ m)数量占总淀粉数的百分比分别为92.14%~94.11%和5.89%~7.86%; 大淀粉粒(粒径> 20 μ m)所占比例微乎其微, 这表明水稻胚乳淀粉几乎由小淀粉粒组成。成熟期水稻强、弱势粒间的小淀粉粒和中淀粉粒数量比例差异不明显, 大淀粉粒虽然数量极少, 但表现出弱势粒中多于强势粒(表2)。
| 表2 水稻胚乳淀粉粒的数量、体积和表面积分布比例 Table 2 Proportion of starch granule number, volume, and surface area distributions in rice endosperm (%) |
两供试小麦品种的小淀粉粒(粒径< 5 μ m)和中淀粉粒(粒径为5~50 μ m)的数量百分比分别为99.70%~99.77%和0.23%~0.30%, 大淀粉粒(粒径> 50 μ m)所占比例很少, 这表明小麦胚乳淀粉基本上由小淀粉粒组成。成熟期小麦强、弱势粒间的小淀粉粒和中淀粉粒数量比例差异不明显, 大淀粉粒数量极少, 且弱势粒中多于强势粒(表3)。
| 表3 小麦胚乳淀粉粒的数量、体积和表面积分布比例 Table 3 Proportion of starch granule number, volume, and surface area distributions in wheat endosperm (%) |
两供试玉米品种的小淀粉粒(粒径< 4 μ m)和中淀粉粒(粒径为4~50 μ m)的数量百分比分别为99.69%~99.75%和0.25%~0.31%, 而大淀粉粒(粒径> 50 μ m)所占比例更少。表明玉米胚乳淀粉基本上由小淀粉粒组成。成熟期玉米强、弱势粒间小淀粉粒和中淀粉粒数量比例差异不明显, 而大淀粉粒数量极少, 且在弱势粒中多于强势粒。
两供试水稻品种的小淀粉粒(粒径< 1.5 μ m)、中淀粉粒(粒径为1.5~20 μ m)和大淀粉粒(粒径> 20 μ m)的体积百分比分别为9.01%~10.80%、73.45%~84.08%和5.12%~17.41%, 说明中淀粉粒(粒径为1.5~20 μ m)占淀粉粒总体积的主要部分(表2)。而且强势粒的中淀粉粒所占百分比显著高于弱势粒。武运粳24和扬两优6号强势粒的中淀粉粒所占体积分别比弱势粒高出13.50%和9.30%。与大淀粉粒所占数量百分比一致, 弱势粒的大淀粉粒体积百分比高于强势粒(表2)。
两供试小麦品种的小淀粉粒(粒径< 5 μ m)、中淀粉粒(粒径为5~50 μ m)和大淀粉粒(粒径> 50 μ m)的体积百分比分别为9.11%~10.20%、78.76%~88.74%和1.36%~11.54%, 这表示小麦淀粉粒的总体积主要决定于中淀粉粒(表3)。在供试的2个小麦品种中, 强势粒的中淀粉粒所占比例大于弱势粒。小麦品种扬麦16和宁麦13强势粒的中淀粉粒所占体积分别比弱势粒高出11.62%和5.14%。与此相反, 弱势粒的大淀粉粒所占百分比高于强势粒(表3)。
两供试玉米品种的小淀粉粒(粒径< 4 μ m)、中淀粉粒(粒径为4~50 μ m)和大淀粉粒(粒径> 50 μ m)的体积百分比分别为5.99%~7.42%、75.20%~88.97%和3.61%~18.81% (表4)。与水稻和小麦一致, 玉米淀粉粒的体积主要决定于中淀粉粒, 且强势粒的中淀粉粒所占体积百分比也高于弱势粒, 弱势粒的大淀粉粒所占体积比例高于强势粒(表4)。
| 表4 玉米胚乳淀粉粒的数量、体积和表面积分布比例 Table 4 Proportion of starch granule number, volume, and surface area distributions in maize endosperm (%) |
两供试水稻品种的小淀粉粒(粒径< 1.5 μ m)、中淀粉粒(粒径为1.5~20 μ m)和大淀粉粒(粒径> 20 μ m)的表面积百分比分别为39.43%~40.53%、58.58%~ 60.18%和0.20%~1.32%, 表明淀粉粒表面积主要取决于小淀粉粒和中淀粉粒(表2)。强、弱势粒间比较, 强势粒的中淀粉粒所占表面积百分比高于弱势粒, 弱势粒的大淀粉粒所占表面积百分比高于强势粒(表2)。
两供试小麦品种的小淀粉粒(粒径< 5 μ m)、中淀粉粒(粒径为5~50 μ m)和大淀粉粒(粒径> 50 μ m)的表面积百分比分别为55.47%~58.25%、40.29%~43.84%和0.21%~1.86% (表3)。强势粒的中淀粉粒所占表面积百分比高于弱势粒, 大淀粉粒的表面积比例表现为弱势粒高于强势粒(表3)。
两供试玉米品种的小淀粉粒(粒径< 4 μ m)、中淀粉粒(粒径为4~50 μ m)和大淀粉粒(粒径> 50 μ m)的表面积百分比分别为54.91%~55.12%、43.43%~44.77%和0.31%~1.45% (表4)。与水稻和小麦一样, 2个玉米品种强势粒的中淀粉粒所占表面积百分比高于弱势粒, 大淀粉粒所占比例低于弱势粒(表4)。
据报道, 水稻的淀粉体通常以多个淀粉粒紧密组装的形式存在于胚乳中, 其淀粉体中组成复粒淀粉体的单个淀粉粒起先是圆球体状的, 当多个淀粉粒紧密结合在一起的时候, 接触面处会形成棱角。在淀粉粒提取的过程中, 单个淀粉粒会释放出来, 表现为有棱角的无规则的单个小淀粉粒[23, 24]。小麦胚乳细胞内的淀粉粒大小分明, 较大的淀粉粒呈两面凸状, 似扁豆, 而小型的淀粉粒呈小球状[25]。玉米胚乳中淀粉粒具有大、中、小3种类型[26]。本研究中, 供试3种作物各自籽粒中的淀粉粒形态特征与前人的描述[23, 24, 25, 26]基本一致, 而且淀粉粒平均粒径表现为玉米> 小麦> 水稻, 这与它们的粒重大小相一致。但是在淀粉粒度分布上, 本文的结果与前人的研究[13, 15, 16, 27, 28, 29]存在差异。Dhital等[27]发现水稻淀粉粒的体积分布会因水稻类型(野生型、转基因或突变体)的不同而不同, 可能表现为三峰或四峰的不同分布趋势。本研究中, 供试的2个水稻品种的淀粉粒数量、体积和表面积分布分别为单峰分布、三峰分布和双峰分布。戴忠民等[28]指出, 小麦淀粉粒的数目表现为单峰分布, 体积和表面积均表现为双峰分布。蔡瑞国等发现糯小麦[15]和强筋小麦[29]籽粒淀粉粒度分布特征均与之相一致。余静等[30]在糯小麦陕糯1号和非糯小麦西农1330淀粉粒径的差异比较中发现, 两品种的淀粉粒数量和表面积均分别呈单峰和三峰分布, 但两品种间存在差异, 陕糯1号和西农1330的淀粉粒体积分别表现为四峰和双峰曲线分布。本文中, 2种供试小麦淀粉粒的数量分布与他们的结果均相同, 但体积和表面积却分别呈现微弱的四峰和三峰分布, 与上述糯小麦陕糯1号淀粉粒的粒度分布趋势基本吻合。张丽等[13]报道, 玉米淀粉粒数量、体积和表面积分布分别表现为单峰、双峰和三峰分布。崔丽娜等[16]发现, 玉米籽粒中淀粉粒数量表现为双峰分布, 体积和表面积均表现为三峰分布。石德杨等[31]在玉米淀粉粒体积和表面积分布的研究结果与崔丽娜等[16]的相同, 但他们发现淀粉粒数量分布趋势与张丽等[13]的一致。本文中发现, 供试的2个玉米品种的淀粉粒数量、体积和表面积分布分别呈单峰、三峰和双峰分布。综上, 不同作物间淀粉粒形态和淀粉粒度分布特征均存在差异, 说明不同作物籽粒的淀粉粒形态和淀粉粒度分布具有各自的特异性, 这可能是不同作物籽粒淀粉具有不同用途的内在原因。相同作物的不同品种间淀粉粒度分布趋势也有不同之处, 表明不同品种籽粒淀粉粒度分布具有个体性。另外, 前人认为作物籽粒中最大的淀粉粒粒径在50 μ m左右 [13, 15, 16, 27, 28, 29], 本文中发现了在50~150 μ m的大型淀粉粒, 但数量极少。近年来, Dhital等 [27]、余静等[30]和崔丽娜等[32]分别在水稻、糯小麦和玉米淀粉粒研究中, 也发现了在50~150 μ m粒径范围内的大型或超大型淀粉粒, 其数量也较少。说明作物籽粒中50~150 μ m的大型或超大型淀粉粒是确实存在的。
本研究中发现, 相同作物强、弱势粒在淀粉粒形态和淀粉粒度分布趋势方面差异并不显著(图1~图4)。但淀粉粒在不同粒径分布比例上存在差异。按不同作物各自的淀粉粒体积分布特点, 将淀粉粒划分为小淀粉粒、中淀粉粒和大淀粉粒(表2~表4)。经比较, 3种作物的强、弱势粒间小淀粉粒粒度分布比例及中淀粉粒所占数量比例没有明显差异, 但各作物强势粒的中淀粉粒所占的体积和表面积比例均显著高于弱势粒, 大淀粉粒的分布比例低于弱势粒。前人对作物强、弱势粒间淀粉粒的研究虽然相对较少。但戴忠民等[17]指出, 小麦强、弱势粒间淀粉粒体积和表面积分布具有明显差异, 强势粒的均呈三峰分布, 弱势粒的均为双峰分布。本文中的结果与之不同, 可能是因为选用品种不同或仪器的分析方法存在差异。Li等[5]指出, 小麦穗中部籽粒中A型淀粉粒和B型淀粉粒在数量比例上与穗基部及穗上部籽粒差异并不显著, 然而它们在体积分布比例上分别明显地低于和高于穗基部及穗上部籽粒。陆大雷等[14]报道, 甜玉米、普通玉米和糯玉米籽粒淀粉粒体积分布在果穗不同部位间差异显著。他们发现普通玉米籽粒中粒径为5~17 μ m的淀粉粒所占体积比例表现为中部> 上部> 基部, > 17 μ m的趋势与之相反 [14]。本研究中发现, 供试的3种作物中籽粒的中淀粉粒占淀粉粒总体积的主要部分。而且籽粒的中淀粉粒所占体积百分比均表现为强势粒的明显大于弱势粒, 这与其淀粉积累量和粒重高低相一致。与此同时, 大淀粉粒的体积分布比例均表现为强势粒的低于弱势粒, 这与其淀粉积累量和粒重高低正好相反。近年, 作者也以盆栽种植的方式, 研究了不同土壤落干对小麦强、弱势粒的淀粉粒度分布的影响。结果表明, 与正常灌溉相比, 土壤轻度干旱增加了弱势粒粒重, 进而提高产量; 土壤重度干旱均降低了强、弱势籽粒重。与此同时, 轻度干旱显著增加了弱势粒的中淀粉粒体积百分比, 降低了弱势粒的大淀粉粒比例; 土壤重度干旱均降低了强、弱势粒的中淀粉粒体积百分比和增加了大淀粉粒的比例(数据未列出)。说明较高的中淀粉粒的体积百分比和较低的大淀粉粒所占比例可能是强势粒具有较高粒重的原因。表明淀粉粒体积是决定粒重高低的一个重要因素, 增加弱势粒的中淀粉粒体积或减小大淀粉粒体积可望增加其粒重。
有研究报道, 在禾谷类作物胚乳发育过程中, 大淀粉粒的发育形成一般较早, 小淀粉粒的发育形成相对较晚[33, 34, 35]。因强势粒胚乳淀粉体发育启始时间通常要早于弱势粒, 故强势粒大淀粉粒所占比例明显高于弱势粒。但本研究观察到, 弱势粒大淀粉粒(粒径> 50 μ m)比例高于强势粒。本研究结果与前人研究结果的差异, 可能与不同类型淀粉粒的粒径划分标准以及所用的品种不同有关。蔡瑞国等[29]和谭秀山等[35]将小麦淀粉粒分为A型淀粉粒(粒径10~45 μ m的大型淀粉粒)、B L型淀粉粒(粒径为1~10 μ m的B型淀粉粒)和B S型淀粉粒(粒径为< 1 μ m的B型淀粉粒)。谭秀山等[35]曾研究了小麦不同穗位强、弱势粒的淀粉粒度分布特性。其结果显示, 麦穗中部的强势粒中A型淀粉粒(粒径为10~45 μ m)所占体积和表面积比例均显著高于麦穗上部或下部的弱势粒。这与本研究的结果基本一致。以往的研究还表明, 强、弱势粒的大淀粉粒所占比例因品种不同而有较大差异 [14, 17, 35]。但对于强、弱势粒大淀粉粒所占比例在不同品种间差异的原因, 尚需深入研究。
又有学者提出, 淀粉分支酶(SBE)不易与小麦B型淀粉粒结合, 但容易与小麦A型淀粉粒结合[36]。伊祖涛等[37]的研究指出, 糯玉米籽粒灌浆过程中, 可溶性淀粉合酶(SSS)和淀粉分支酶(SBE)活性与大淀粉粒(粒径> 7.4 μ m)的体积增大速率和平均粒径增大速率呈显著或极显著正相关, SSS和SBE活性越高, 淀粉粒平均粒径和大淀粉粒(粒径> 7.4 μ m)体积分布比例就越大。Zhang等[33]提出小麦强、弱势粒中的淀粉粒度分布的不同特性与其灌浆进程中的淀粉合成相关酶活性及其基因表达的差异密切相关。McMaugh等[38]利用RNA干扰技术人为抑制淀粉合酶I (SSI)在小麦胚乳发育中的表达, 结果表明, 直/支链淀粉的精细结构与含量及其淀粉粒的形态结构、分布比例均发生了显著变化, 进而改变了淀粉的品质特性(例如:糊化温度、膨胀势和黏度特性等)。说明SSI在决定胚乳淀粉结构与特性方面发挥着重要作用。作者据此推测, 籽粒灌浆过程中淀粉合成相关酶活性或基因表达的差异可能是导致强、弱势粒间淀粉粒度分布比例差异的原因, 其机制有待深入研究。
籽粒的淀粉粒形态大小在水稻、小麦和玉米3种不同作物间存在明显差异, 粒径上表现为玉米> 小麦> 水稻。各作物按粒径可将籽粒淀粉粒分为小淀粉粒、中淀粉粒和大淀粉粒3种类型。籽粒淀粉粒的总体积主要决定于中淀粉粒体积。各作物强势粒的中淀粉粒所占的体积百分比均显著高于弱势粒, 大淀粉粒所占的体积百分比则显著低于弱势粒。强、弱势粒的中淀粉粒所占体积比例与其淀粉积累量和粒重变化趋势一致。增加弱势粒的中淀粉粒体积或减小大淀粉粒体积可望增加其粒重。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|