

{kind=link}
{kind=link}
{kind=link}
{kind=link}
一个新的水稻内卷叶突变体的表型和遗传分析
[解志伟1, 2 , 孙伟3 , 尹亮3 , 赵金凤2 , 袁守江3 , 张文会1, *  , 李学勇
, 李学勇2, * ]
, 李学勇]
|
|


叶片是光合作用的重要器官, 适度卷曲有利于改善群体光照、提高光能利用率, 卷叶基因是培育理想株型的重要资源。为研究控制水稻叶片形态建成的分子机制, 从EMS诱变粳稻品种日本晴的M2代中分离了一个叶片向内卷曲的突变体
, LI Xue-Yong
Leaf is an important organ for photosynthesis. Moderate rolling of leaves can facilitate the improvement of plant’s population structure and enhance light-use efficiency, which is very important in ideotype breeding of rice. In the present study, in order to systematically dissect the molecular mechanism of leaf morphogenesis and development, one ethyl methylsulfone (EMS) -induced rice (
叶片是光合作用的主要器官, 适度卷曲的叶片不但能够很好地保持直立状态, 改善其受光姿态, 提高光能利用效率, 还可以减少叶片蒸腾, 提高植株对干旱环境的耐受能力, 有利于提高产量和品质[1,2,3,4]。因此, 叶片形态改良一直是株型育种的重要目标, 卷叶品种或突变体是遗传与育种研究的重要资源。
已发现的水稻卷叶突变体主要有两类, 一类的叶片表现近轴化发育, 向内卷曲, 目前已报道的有 B157[5] 、rl(t)[6] 、sll1(rl9)[7,8] 、nrl1[9]等; 另一类的叶片表现远轴化发育, 向外卷曲, 如 oul1[10] 、adl1[11]和 acl1[12]等。玉米、金鱼草( Antirrhinum majus )等植物叶片的发育研究表明, 控制叶片近轴/远轴发育的有 PHAN[13]、 HD-ZIPIII[14]、 YABBY[15]和 KANADI 等基因家族中的基因[16]。近年来, 已分离出几个水稻卷叶基因。如Yan等[8]分离的卷叶基因 RL9 , 认为可控制叶片的背腹极性, 最终引起叶片卷曲。Zhang等[7]随后分析了 RL9 的等位基因 SLL1 , 发现该基因编码的MYB转录因子与拟南芥 KANADIs 基因家族同源, 其功能缺失会导致远轴叶肉细胞的程序化死亡, 抑制远轴特征的发育, 叶片上表面的泡状细胞的数目和大小都有不同程度的减少, 从而引起 sll1 突变体叶片高度内卷。Shi等[17]分离出一个与拟南芥 ZIP/Ago7 同源的 OsAGO7 基因, 该基因的过量表达同样会导致叶片上表面泡状细胞数目和大小的减少, 导致叶片向内卷曲。Fang等[18]分离出一个内卷叶突变体 rl14 , 与野生型相比 , 表现气孔复合体变小, 蒸腾速率下降, 叶片上表面泡状细胞体积明显收缩。 RL14 基因通过影响次生壁的形成来影响水分的运输, 内卷叶突变体 rl14 泡状细胞形态的异常主要是水分运输改变而引起的缺水所致。Xiang等[19]分离了一个卷叶基因 SEMI-ROLLED LEAF1(SRL1) , 可通过负调控编码H+-ATP酶亚基和H+-焦磷酸酶的基因的表达来抑制泡状细胞的形成, 最终调节叶片的卷曲。此外, Zou等[10]通过T-DNA插入的方法获得一个外卷叶突变体 oul1 , 其叶片外卷是敲除水稻 ROC5 基因造成的。
本研究利用EMS (ethylmethane sulfonate)化学诱变日本晴获得内卷叶突变体 s1-145 , 对其进行了表型鉴定、遗传分析和基因定位, 为进一步克隆该基因和解析控制叶片卷曲的分子机制提供了资源和技术支持。
用EMS化学诱变粳稻品种日本晴, 获得了内卷叶突变体 s1-145, 多年连续种植, 卷叶突变性状能够稳定遗传。遗传分析和基因定位所用的父本为籼稻品种Dular, 整个生育期叶片都表现平展。
1.2.1 突变体的表型鉴定 在孕穗期随机选取 s1-145 突变体与野生型日本晴各15株, 测定其剑叶长度并于叶片长度1/2处测量叶宽。同时计算剑叶的叶片卷曲指数LRI (leaf rolling index)[20], LRI = ( Lw - Ln) / Lw。其中 Lw为叶片展开时最宽处的宽度(cm), Ln为叶片最宽处卷曲后两叶缘之间的距离(cm)。
1.2.2 遗传分析和F2定位群体构建 2009年夏用卷叶突变体 s1-145 与广亲和籼稻Dular杂交, 同年冬季在海南种植Fl并收获F2种子。2010年夏在山东省水稻研究所种植F2群体, 观察各株表型, 选取卷叶单株作为定位群体。
1.2.3 叶绿素含量和光合速率与蒸腾速率的测定
在抽穗期分别选取卷叶突变体 s1-145 和日本晴各10株, 取其剑叶最宽处测定叶绿素含量[21]。利用LI-6400XT便携式光合测定仪分别测定剑叶最宽处的光合速率与蒸腾速率。
1.2.4 水稻叶片DNA的提取与基因初步定位 采用改进的CTAB (cetyltrimethyl ammonium bromide)法提取亲本及定位群体的基因组DNA[22]。参照郭伟伟等的方法[22]用混合分离群体分析法(bulked segregant analysis, BSA)构建混池。调节2个亲本DNA浓度使其一致, 等量混合亲本基因组DNA, 构建亲本混池; 选取浓度相近的10株定位群体单株的基因组DNA, 等量混合构建定位群体混池。
选取本实验室已发展的平均覆盖水稻12条染色体的163对InDel (insertion-deletion)标记对突变体及2个亲本进行BSA分析, 找出可能与卷叶基因连锁的标记, 然后用这些可能连锁的标记引物对F2群体进行单株验证。根据初步定位的结果以及比对日本晴和Dular在该区间的序列, 利用DNAStar软件设计新的InDel引物(表1)。PCR总体系为10 μL, 包含2.0 μL 5 × Mix buffer、6.0 μL ddH2O、1.0 μL 2.0 μmol L-1引物、1.0 μL模板DNA。PCR程序为94℃预变性3 min; 94℃变性30 s, 57℃退火30 s, 72℃复性30 s, 共35个循环; 72℃延伸10 min。将PCR产物在8%的非变性聚丙烯酰胺凝胶上电泳, 快速银染后观察。统计每个F2突变个体的基因型, 利用MAPMAKER/EXP3.0软件计算定位标记与目标基因之间的遗传距离。
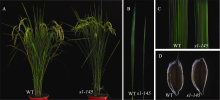
在EMS诱变粳稻品种日本晴的M2代中选出1株内卷叶突变体 s1-145 , 其植株直立紧凑(图1-A)、叶片变窄(图1-C)、叶缘沿中轴内卷形成圆筒状(图1-B)、植株高度无明显变化(图1-A), 生育期稍延迟(图1-A), 育性和粒形不变(图1-D)。 s1-145 的叶长与野生型差异不明显(图2-A), 但叶片宽度明显变窄, 只有野生型的60%左右(图2-B)。
水稻叶片的卷曲与否以及卷曲程度可以用叶片卷曲指数(leaf rolling index, LRI)衡量。 LRI = 0时, 叶片平展; LRI ≥1时, 叶片极度卷曲。卷叶突变体 s1-145 的叶片卷曲指数在0.6左右, 而野生型叶片的LRI值接近0 (图2-C)。
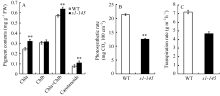
与已报道的内卷叶突变体 sll1 相似[7], s1-145 叶色比野生型更加深绿。 s1-145 叶绿素a和总叶绿素(a + b)含量均显著高于野生型日本晴(图3-A)。另外, 突变体类胡萝卜素含量也显著高于日本晴(图3-A)。
 | 图 1 卷叶突变体 s1-145 与野生型日本晴的形态比较A: 灌浆期株型; B: 剑叶(自然状态); C: 剑叶中部(平展状态); D: 粒形。Fig . 1 Morphology of the s1-145 mutant and wild type NipponbareA: plant architecture at filling stage; B: flag leaf (natural state); C: central part of flag leaf (flattened state); D: grain shape. |
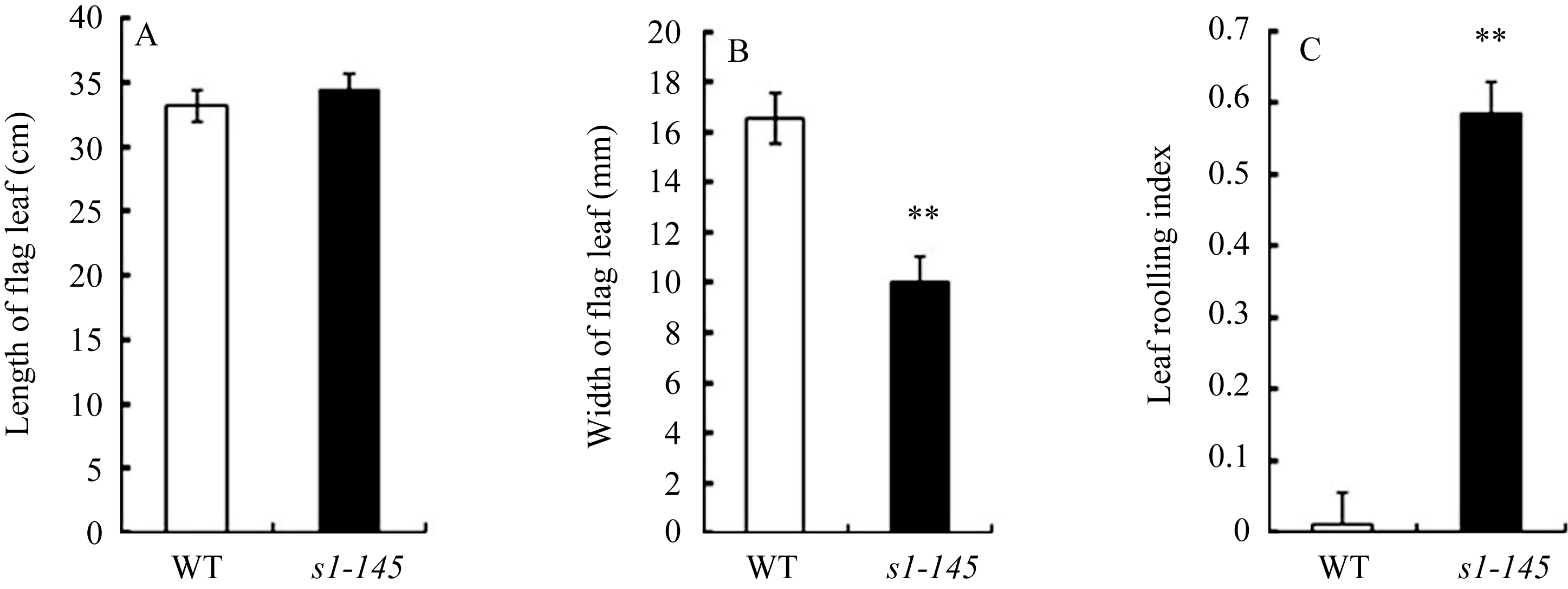 | 图 2 卷叶突变体 s1-145 与野生型日本晴的叶片形态指标A: 剑叶长度; B: 剑叶宽度; C: 叶片卷曲指数。数值=平均值±标准误差, n = 10。** 表示野生型与突变体相比在 P<0.01水平差异显著。Fig . 2 Leaf morphological parameters of the s 1-145 mutant and wild type NipponbareA: Length of the flag leaf; B: Width of the flag leaf; C: Leaf rolling index. Value = mean ± SE, n = 10. ** Represents significantly different at P< 0.01. |
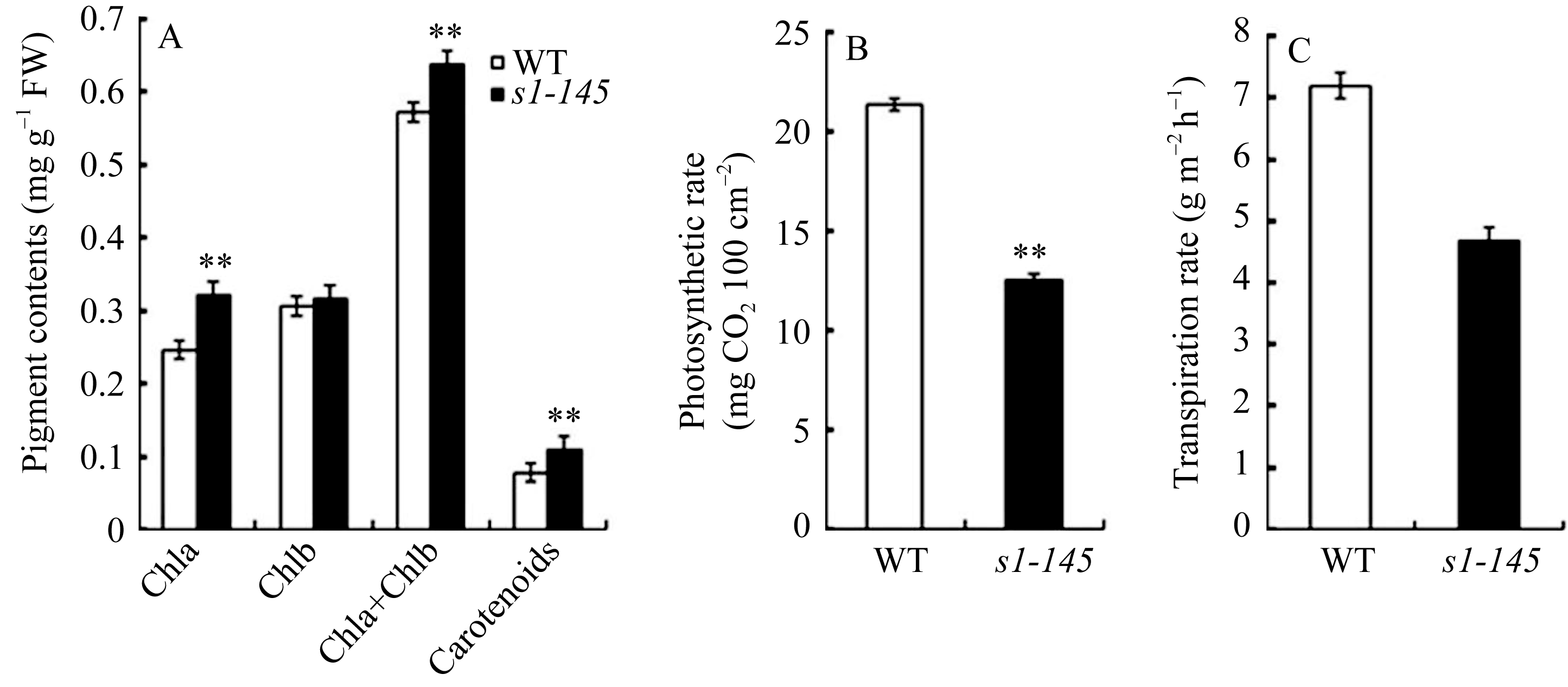 | 图 3 卷叶突变体 s1-145 和野生型剑叶生理指标A: s1-145 和野生型剑叶叶绿素含量; B和C: s1-145 和野生型剑叶的光合速率和蒸腾速率。数值=平均值±标准误差, n = 5。**表示野生型与突变体相比在 P<0.01水平差异显著。Fig . 3 Physiological parameters of the flag leaf in wild type and s1-145 mutantA: chlorophyll content of the s1-145 and WT flag leaves; B and C: photosynthetic rate and transpiration rate of the wild type and s1-145 mutant flag leaves. Value = mean ± SE, n = 5. **Represents significantly different at P< 0.01. |
植物叶片形态及叶绿素含量的改变势必影响叶片的光合效率。卷叶突变体 s1-145 单个剑叶的光合速率低于野生型日本晴(图3-B)。这可能是叶片内卷导致受光面积和强度降低所致, 但这并不意味着田间群体的光合效率会降低, 因为适度卷曲的叶片能够保持直立状态, 有利于改善群体光照进而提高光能利用率。突变体 s1-145 的蒸腾速率也低于野生型日本晴(图3-C), 这说明叶片卷曲使水分丧失变慢, 可能有助于提高水稻的耐旱性。
用纯合内卷叶突变体 s1-145 与叶片平展的广亲合籼稻品种Dular杂交, F1植株均表现正常, F2群体出现性状分离。在调查的1338株F2单株中正常叶植株1037株, 卷叶突变体植株311株, 经卡方检测(χ2= 1.32< 3.84 [χ2(0.05,1)]), 正常和突变表型个体的分离比例符合3∶1, 推断该突变性状受1对隐性基因控制。
采用混合分离群体分组分析法(BSA)对F2卷叶个体混池和平展叶个体混池, 利用本实验室开发的覆盖水稻全基因组的163对InDel标记进行扫描。发现位于第2染色体的2个InDel标记R2-33.26和R2-16可能与目标基因连锁。随后用24株F2突变个体单株验证表明, 这2个标记均与 s1-145 突变体卷叶基因连锁(图4)。
为了对该卷叶基因进一步精细定位, 在R2- 33.26和R2-16之间又开发了6个新的InDel标记(图4和表1)。利用这些标记对311株F2群体卷叶个体进行连锁分析, 最后将该基因定位在R2-34.70和R2-34.79之间, 遗传距离为0.64 cM, 物理距离约为90 kb (图4)。
| 表 1 本研究所用的引物 Tab le 1 Primers used in this study |
 | 图 4 内卷叶突变体 s1-145 的基因定位图谱上排为初步定位结果, 下排为精细定位结果。横线上方为定位标记, 横线下方数字代表交换单株个数和遗传距离。Fig . 4 Gene mapping of the adaxially-rolled leaf mutant s1-14 5The upper row shows the primary mapping result, while the lower is the fine mapping result. The characters above the line represent the markers used, while the numbers below the line represent the number of recombinants and genetic distance, respectively. |
从表面上看, 叶片仅仅是由几种不同类型的细胞构成, 然而其发育却是一个相当复杂的过程[23,24], 涉及到很多因素, 通过分离鉴定不同的卷叶突变体并克隆基因, 对揭示植物叶片生长发育的分子机制、合理利用有利等位突变基因资源来提高作物产量具有重要意义。已报道的水稻内卷叶突变体 sll1 和 nrl1 都表现育性降低, nrl1 植株还有不同程度的矮化[7,9]。而本文报道的内卷叶突变体 s1-145 育性正常, 植株高度没有明显变化, 株型紧凑, 可以作为育种材料加以利用。在水稻第2染色体长臂上已定位了2个外卷叶突变体即 oul1[10]和 adl1[11], 与本研究的突变体的叶片卷曲方向相反。 oul1 是通过T-DNA插入的方法获得的, 其叶片外卷是敲除水稻 outermost cell-specific gene5 ( ROC5 )基因造成的, ROC5 基因与拟南芥 HD-ZIP IV 基因家族的 GLABRA2 基因同源, ROC5 基因通过负调控泡状细胞的发育来控制叶片卷曲。与野生型相比 oul1 突变体中的泡状细胞体积变大, 数目增多, 导致叶片向外卷曲。过表达 ROC5 基因, 叶片上表面叶脉两侧泡状细胞数目减少, 体积变小, 导致叶片内卷。 adl1 叶片上表面由泡状类细胞覆盖, adl1 突变体中 HD-ZIP III 基因在幼嫩的叶原基没有表达, 但是在成熟的叶片中表达量增多, ADL1 编码植物特有的类半胱氨酸钙蛋白酶, 与玉米的 DEFECTIVE KERNEL1 同源, ADL1 不直接参与初始叶极性的发育, 而是参与叶轴极性的构建, 推测 ADL1 通过促进适当的表皮细胞的发育来影响叶片和胚的发育模式。 s1-145 虽然也定位在第2染色体长臂, 但与 oul1 和 adl1 不在同一定位区间内。
根据美国水稻基因组注释项目预测(http://rice. plantbiology.msu.edu/), 查阅第2染色体上目标区段内所有的预测基因, 发现这13个预测基因中, 编码假定蛋白的基因2个; 反转座子基因4个; 编码功能蛋白的基因7个, 在这7个有功能注释的基因中有1个编码水稻WAK类受体蛋白激酶, 2个基因编码脱氢酶, 1个编码F-box结构域蛋白, 1个基因编码还原酶, 1个编码翻译起始因子eIF-2B亚基, 还有1个编码肉桂酰辅酶A还原酶, 但没有发现与已报道的控制叶片近轴化发育的基因或同源基因。因此, s1-145 是一个新型内卷叶突变体。为了获得控制 s1-145 叶片卷曲的基因, 需要进一步开展精细定位和基因克隆等工作。
以EMS化学诱变从粳稻品种日本晴获得了一个内卷叶突变体 s1-145 , 叶片内卷并且变窄, 生育期稍延迟, 株高和育性等农艺性状正常。该性状由1对隐性基因控制, 被定位到第2染色体长臂上90 kb区间内。 s1-145 是一个新型内卷叶突变体, 为水稻株型改良提供了材料和基因资源。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|