
 stratification
stratification ), and germination of conifer seeds is discussed.
), and germination of conifer seeds is discussed.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
种子萌发及其调控的研究进展
[徐恒恒1  , 黎妮
, 黎妮2 , 刘树君1 , 王伟青1 , 王伟平2 , 张红1 , 程红焱1 , 宋松泉1, * ]
, 黎妮]
|
|


第一作者联系方式: E-mail:Hengheng0425@126.com
大多数有花植物通过有性生殖和产生种子繁衍后代, 种子的成功萌发和正常成苗决定植物物种的繁衍与生存。种子萌发容易受到机械伤害、病害和环境胁迫的影响, 是植物生活周期中最重要和最脆弱的阶段, 对于一年生和二年生植物则更为重要。种子萌发是一个复杂的多步骤过程, 在这个过程中静止的干燥种子迅速恢复代谢活性, 完成胚伸出周围结构的细胞事件, 以及为随后的幼苗生长做准备。本文综述了近年来种子萌发及其调控的研究进展, 主要包括种子萌发过程中的重要生理事件, 与种子萌发有关的蛋白合成、翻译后修饰和蛋白质组, 以及植物激素对种子萌发的调节。此外, 我们还提出了种子萌发的能量刺激假说, 此假说为减少禾谷类作物种子和粮食生产中发生的穗萌现象提供了新的研究思想, 并且有助于回答植物学、农学和园艺学中的2个基本问题, 即胚怎样从它的周围结构中伸出完成萌发?胚的伸出怎样被阻断以至于种子被维持在休眠状态?
, LI Ni
Most flowering plants reproduce through sexual breeding and seed production. The success of seed germination and the establishment of a normal seedling determinate the propagation and survival of plant species. Seed germination is the most critical and flimsy phase in plant life cycle because of its high vulnerability to injury, disease and environmental stress, which are very important especially for annual and biannual plant species. Seed germination is a complex and multistep process during which the quiescent dry seeds quickly recover the metabolic activity, complete essential cellular events for the embryo to emerge from surrounding structures, and prepare for subsequent seedling growth. In the present paper, the recent research progress in seed germination and its control is reviewed, mainly including the important physiological events during germination, the protein synthesis, posttranslational modification and proteome associated with seed germination, and regulation of phytohormone on seed germination. Furthermore, we have also proposed an energy stimulating hypothesis for seed germination, which provides a new idea for decreasing the pre-harvest sprouting occurred in seed and foodstuff production of cereal crops and answering two fundamental questions in botany, agronomy, and horticulture: how does the embryo emerge from its surrounding structures to complete germination and how is embryo emergence blocked so that seeds can be maintained in the dormant state?
大多数有花植物通过有性生殖和产生种子繁衍后代, 种子活力的保持和成功萌发成苗决定着植物种群的繁衍和生存[ 1], 也决定植物进入自然和农业生态系统的时间, 直接影响作物的产量[ 2]。因此, 种子萌发具有重要的经济和生态意义。由于种子萌发容易受到机械伤害、病害和环境胁迫的影响, 种子萌发也被认为是植物生活周期中最重要和最脆弱的阶段[ 1]; 对于一年生和二年生植物, 种子的成功萌发则更为重要。
种子萌发起始于水分吸收(吸胀), 结束于胚轴的伸出, 通常是胚根突破周围结构[ 3]。胚轴突破周围结构也被称为“可见萌发”, 此时, 种子已经完成萌发。萌发(germination)的术语通常被用来表示它的完成, 例如, “50%的萌发”表示一个种子批中50%的种子已经萌发。狭义的萌发( sensu stricto germination)不包括幼苗生长, “萌发的幼苗(germinating seedling)” 的术语具有明显的错误[ 3]。萌发有时也被种子检验员称为从土壤中出苗, 因为他们的兴趣在于检测幼苗的建成。一个更为准确的术语是“出苗(seedling emergence”或者是“成苗(seedling establishment)”。在新生幼苗中发生的事件, 例如主要贮藏物的动员, 也不属于萌发的内容, 它们是萌发后的事件。Bewley等[ 3]注意到有大量关于种子生理、细胞学和分子生物学的文献, 是利用已萌发的种子(或者幼苗)作材料来研究种子萌发的机制, 错误地描述种子萌发的结论; 因此, 从这些文献中获取信息时必须小心。
在许多栽培作物特别是一些重要的禾谷类作物, 例如水稻、小麦和玉米中, 由于长期的栽培选择, 即种子迅速和整齐的萌发, 使其种子在成熟后期遇到连续阴雨时会产生收获前萌发, 也称穗萌(pre- harvesting sprouting)[ 4, 5]。穗萌是种子在植株上的一种特殊萌发, 在禾谷类作物种子和粮食生产中已经造成巨大的经济损失[ 4], 是目前全球关注的重要问题。
Rajjou等[ 1]提示, 从1900年以来, 发表了25 000多篇关于种子萌发的文献。这些文献充分表明了种子萌发在农业、林业和园艺生产中的重要性以及在不同物种中的复杂性。Nonogaki等[ 6]认为, 尽管获得了种子萌发过程中一些基因表达的信息, 但由于种子萌发事件的复杂性以及不同植物种子的遗传多样性, 完成种子萌发的关键事件仍未确定。本文综述了近年来种子萌发及其调控的研究进展, 主要包括种子萌发过程中的重要生理变化, 与种子萌发有关的蛋白质合成、翻译后修饰和蛋白质组, 以及种子萌发的激素调节, 试图为解释种子萌发的机制, 促进引种时种子能迅速而整齐地萌发并减轻或防止收获前穗萌提供新的研究思想。
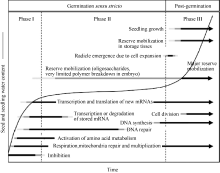
当具有生活力的干燥种子吸收水分时, 一系列的事件被启动, 最终引起胚通常是胚根的伸出, 表明种子已成功地完成萌发。种子萌发过程中发生的细胞变化是复杂的, 当吸胀时, 代谢活性迅速增加, 由成熟干燥和干燥种子时氧化所引起的结构伤害被恢复, 基本的细胞活性被重新激活, 胚为出苗和随后的早期幼苗生长作好准备(图1)[ 3]。
种子萌发通常被描述为一个由3个阶段所组成的过程, 随着水分吸收开始(阶段I, 吸胀作用), 重新开启代谢过程(阶段II), 胚根突破周围结构完成萌发(阶段III) (图1)[ 3]。干燥种子具有非常低的水势[ 7], 能使水分迅速地进入。由于这个过程被衬质势(matrix potential)所驱动, 吸胀作用也在死种子中发生[ 8]。水分吸收的动力学受种子结构的影响, 水分不可能均等地进入种子的所有部位[ 9]。最初, 吸胀作用引起衬质例如细胞壁和细胞内的贮藏多聚物的水合。水分吸收随着时间的延长而增加是种子内水合细胞数量的增加, 而不是所有细胞水合作用一致增加的结果。利用质子核磁共振微成像(proton nuclear resonance micro-imaging)技术能够检测到吸胀过程中的水合模式。在烟草种子中, 水分首先到达珠孔端胚乳和胚根的顶部[ 10], 使该区域的代谢作用迅速恢复。许多其他的植物种子包括小麦种子也是通过珠孔发生类似的水分吸收模式[ 11]; 但也有例外, 例如西部白松, 种子合点端的子叶是最先水合的部位[ 12]。在吸胀过程中, 种子迅速膨胀, 其大小和形状也发生变化。Robert等[ 13]发现, 乙烯突变体的种子形状和吸胀行为不同于野生型种子。吸胀种子的大小和形状变化差异可用于大规模的突变体筛选, 计算方法促进了图像时间序列(image time series)的高通量分析[ 14]。
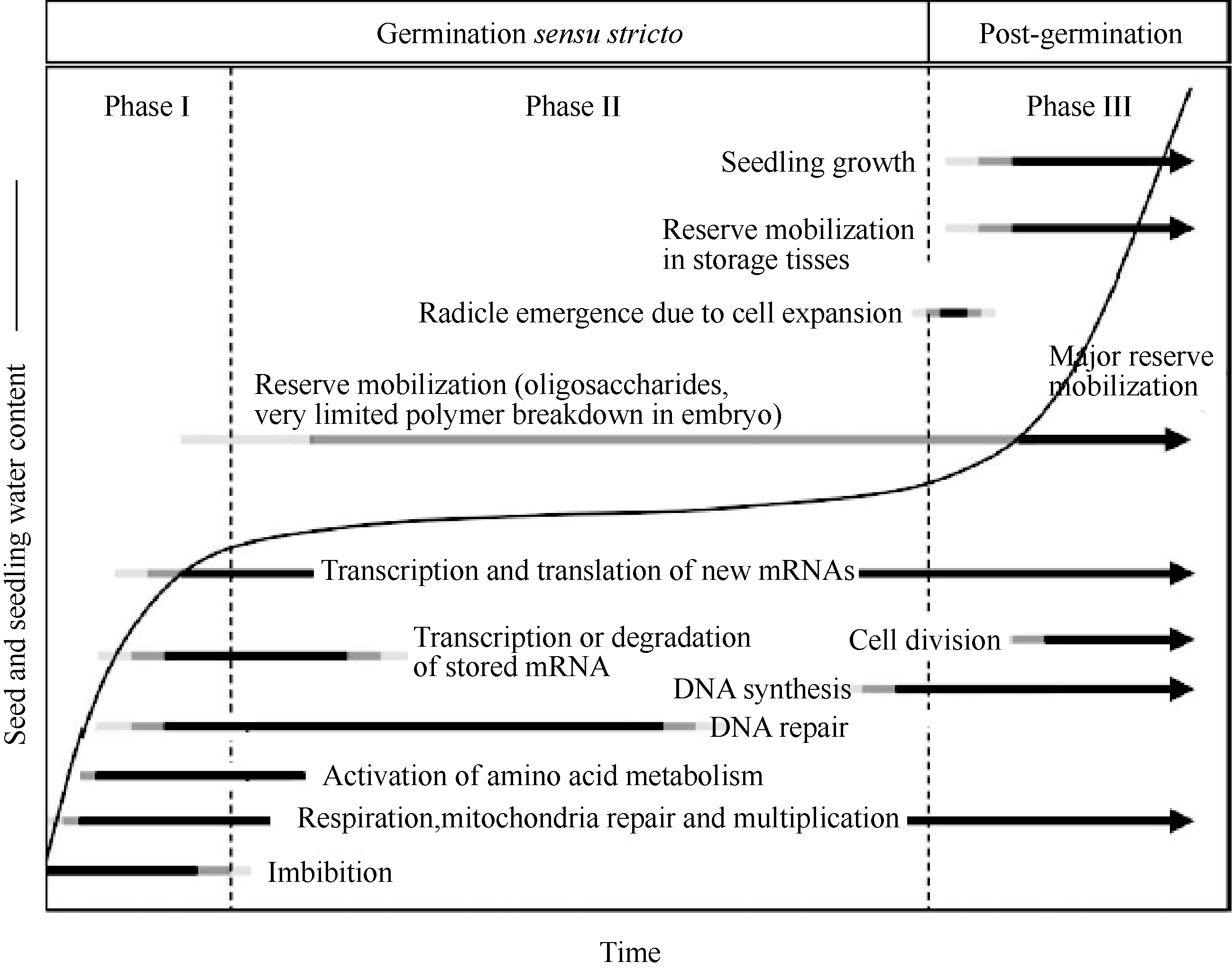 | 图1 水分吸收的时间进程以及一些重要的与萌发和早期幼苗生长有关的变化。Fig. 1 Time course of water uptake and some important changes associated with germination and early seedling growth |
种子的最初吸胀作用常常伴随着细胞溶质的大量渗漏。渗漏是由快速和/或者不均匀的重新水合作用所引起的膜和细胞分室作用被伤害的结果。在种子成熟干燥和随后的贮藏过程中, 细胞膜的结构被维持在凝胶相; 当重新水合时, 转变成为液晶相; 在相变过程中, 一些膜被伤害, 导致溶质从细胞内渗漏[ 3]。渗漏也能降低种子内抑制物的浓度, 从而促进种子萌发[ 3]。
在种子吸胀开始后的几分钟, O2的吸收和CO2的释放迅速增加[ 2, 3]。成熟的干燥种子含有内膜分化不完全的线粒体以及呼吸作用必需的功能酶例如三羧酸循环的酶和末端氧化酶, 在吸胀开始后的几小时能通过氧化磷酸化提供足够的ATP[ 6]。在干燥种子中, 这些酶可能被线粒体专一的胚胎发生晚期丰富(late embryogenesis abundant, LEA)蛋白保护[ 15, 16]。在吸胀完成后(时期II), 当线粒体经过修复和复制时, 线粒体的呼吸作用增加。在种子中线粒体有两种不同的发育模式。在贮藏淀粉为主的种子例如豌豆、绿豆和豇豆中, 以预存的线粒体被修复和重新活化为主; 而在贮藏油为主的种子例如花生、蓖麻和南瓜中, 新线粒体的生物发生是主要的[ 6]。
糖酵解和戊糖磷酸途径也在吸胀的种子中具有活性。在吸胀过程中或者吸胀后, 许多种子经历短暂的厌氧条件, 导致乙醇的产生。当线粒体ATP的产生受到低氧限制时(常常是由于胚周围结构的限制), 糖酵解途径占优势; 相反, 当线粒体变得活跃时, 戊糖磷酸途径占优势[ 6]。最近, 我们发现杨树种子在吸胀过程中的呼吸速率迅速增加, 吸胀0.5 h时的种子呼吸速率是干燥种子的5倍; 细胞色素c氧化酶途径抑制剂KCN, 交替氧化酶途径抑制剂水杨基羟肟酸(salicylhydroxamic acid)和解偶联剂2,4-二硝基酚(2,4-dinitrophenol)显著地抑制杨树种子的萌发(宋松泉等, 未发表数据), 表明呼吸作用(能量产生)在种子萌发中起重要作用, 但作用方式还不清楚。
基因组DNA的损伤包括在延长干藏过程中端粒顺序的逐渐丧失[ 17], 链断裂, 以及由温度、水分、氧气和活性氧(reactive oxygen species, ROS)积累所引起的其他类型的DNA损伤[ 18]。染色体损伤的积累和/或者这种伤害在种子吸胀过程中不能被修复似乎是种子贮藏过程中生活力丧失的重要因素。玉米种子成熟过程中的脱水和吸胀过程中的重新水合导致大量的单链DNA断裂, 其中大部分伤害可能归因于吸胀伤害。这包括萌发早期DNA中缺嘌呤/缺嘧啶位点(apurinic/apyrimidinic site)的产生和修复。DNA损伤明显地是种子萌发过程中的一种主要障碍, 能被DNA连接酶(DNA ligase)修复。当拟南芥种子吸胀时, DNA连接酶的表达被迅速活化; 在核DNA复制或者细胞分裂缺乏时, 观察到高水平的DNA重新合成, 表明在DNA修复中的作用[ 2]。在拟南芥中, 植物专一的DNA连接酶VI的失活使种子萌发延迟[ 19]。 atlig6突变体对种子老化也表现出过敏性反应, 表明AtLIG6是拟南芥种子质量和寿命的主要决定子[ 19]。在苜蓿种子萌发的早期阶段, 与氧化损伤的DNA修复有关的酶[例如, 甲酰嘧啶-DNA糖基化酶(forma-midopyrimidine-DNA glycosylase)和8-氧鸟嘌呤DNA糖基化酶/裂解酶(8-oxoguanine DNA glycosylase/ lyase)]被上调也支持DNA的修复是种子萌发所需要的[ 20]。烟碱是多聚腺苷二磷酸-核糖聚合酶[poly (ADP-ribose) polymerase, PARP; 与DNA的修复有关]的抑制剂, 在种子萌发过程中这个化合物必须被降解。在拟南芥成熟种子中, 编码烟酰胺酶(nicotina-midase)的基因 NIC2被高水平地表达[ 21]。敲除突变体 nic2- 1的种子表现出烟酰胺酶活性降低和延迟萌发, 说明烟酰胺被NIC2正常地代谢, 从而释放对PARP活性的抑制, 允许DNA修复发生[ 21]。
在种子干燥和/或者贮藏过程中, 由于异常氨基酸的形成也发生蛋白质的伤害; 这样可能导致蛋白质的错误折叠, 引起蛋白质功能的下降或者丧失。PIMT通过催化异常的L-异天冬氨酰残基转换成为正常的L-天冬氨酰在种子伤害蛋白的修复中起重要作用[ 22, 23]。在诱变处理的拟南芥种子中, PIMT1(编码PIMT的一个基因)的过表达减少蛋白质中异常氨基酸残基的积累, 引起种子活力和寿命的增加; 相反, PIMT1的降低表达增加新鲜收获的成熟干燥种子中蛋白质L-异天冬氨酰残基的积累, 导致胁迫条件下种子活力的下降, 以及对老化处理的过敏性反应[ 23]。PIMT2主要定位于内膜系统, 例如线粒体、叶绿体和细胞质, 提出PIMT2在一些亚细胞中参与蛋白质的修复[ 22]。
在干燥的拟南芥和大麦种子中存在多于12 000种mRNA[ 24, 25], 在干燥的水稻种子中含有超过17 000种mRNA[ 26]。这些mRNA被称为贮存的或者残留的信息, 来源于种子发育, 并在细胞脱水中存留; 但是, 这些信息在干燥种子中的定位是不清楚的, 它们可能被束缚在细胞质内的信使核蛋白复合物(messenger ribonucleoprotein complexes)中[ 6]。在干燥种子中, 例如小麦胚含有蛋白质重新合成所需要的全部组分[ 3]。在重新水合的几分钟内, 能利用存留的mRNA, 核糖体被转变成为多聚核糖体蛋白合成复合物(polysomal protein-synthesizing complexes)[ 3]。转录和翻译抑制剂的实验支持拟南芥种子萌发过程中贮藏mRNA的参与。RNA聚合酶II的抑制剂α-鹅膏蕈碱(α-amanitin)不抑制种子萌发, 而蛋白质合成抑制剂环己酰亚胺(cycloheximide)明显地阻止胚根的伸出[ 27, 28](宋松泉等, 数据待发表)。值得注意的是, 利用存留的mRNA的重新蛋白合成足够允许种子完成萌发, 但是在吸胀早期哪些存留的mRNA是萌发完成所必需的仍然不清楚。
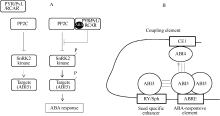
He等[ 29]利用蛋白质组技术构建了萌发水稻种子的代谢和调节途径。拟南芥[ 30, 31, 32]、水稻[ 33, 34]、玉米[ 35]、豌豆[ 36]、番茄[ 37]、蓖麻[ 38]、麻疯树[ 39]和海枣[ 40]种子萌发过程中的差异蛋白质组分析表明, 有许多蛋白被差异积累。这些蛋白主要涉及代谢作用(包括氨基酸、脂类、氮和硫、糖、辅因子和次生代谢), 能量产生(糖酵解、戊糖磷酸途径、三羧酸循环和呼吸作用), 转录, 蛋白质合成与目标(包括蛋白质合成、折叠和水解), 细胞生长与结构(细胞骨架、生长调节), 细胞防御与救援(防御相关蛋白、去毒和胁迫反应)以及贮藏蛋白(图2)。应该注意的是, 由于不同物种的干燥种子中所含的mRNA的种类与丰度不同, 上述蛋白质组研究中所涉及的种子萌发条件不一致, 并且蛋白质组分析所鉴定的蛋白数量有限, 要比较上述物种萌发过程中同一蛋白的积累模式是困难的。
 | 图2 萌发的豌豆种子胚轴中已鉴定的差异积累蛋白Fig. 2 Differentially-accumulated proteins identified in embryonic axis of germinating pea seeds |
根据差异积累的蛋白质组研究, 发现与甲硫氨酸(Met)代谢有关的一些蛋白例如 S-腺苷甲硫氨酸( S-adenosylmethionine, AdoMet)合成酶[ 29, 30, 31, 36, 41, 42]、Met合酶[ 36]、不依赖钴胺素的Met合酶(cobalamin- independent Met synthase)[ 31]、以及半胱氨酸合酶[ 36, 41, 42]在种子萌发过程中增加积累; 这些结果与Rajjou等[ 1]的观点一致, 他们认为Met代谢在种子萌发中起重要作用。在由植物合成的必需氨基酸中, Met是一种重要的代谢物, 因为它不仅是蛋白质合成的底物, 而且是AdoMet (一种普遍的甲基供体)、多胺、植物激素乙烯和维生素生物素的前体[ 43, 44]。已经提出, 在所有的生物中, Met代谢是一种看家机制(housekeeping mechanism)[ 1]。
在拟南芥[ 31]、水稻[ 28, 29, 33, 41, 42]、玉米[ 45]、豌豆[ 36]、麻疯树[ 39]、海枣[ 40]和蓖麻[ 38]种子萌发过程中, 增加积累一些与糖酵解、三羧酸循环、戊糖磷酸途径和能量产生有关的蛋白, 为种子萌发提供能量和碳骨架。在25℃ (萌发)、43℃ (不萌发)、(43+25)℃ (恢复萌发)和ABA (不萌发)中萌发的水稻种子的蛋白质组分析表明, 增加积累蛋白OsJ_19262 (6-磷酸果糖激酶3)和甘油醛-3-磷酸脱氢酶(刘树君等, 未发表数据)。这些结果表明能量供应是种子完成萌发所必需的, 但不是足够的。
与蛋白合成相关的蛋白例如多聚腺苷酸-结合蛋白、真核翻译起始因子5A-3和因子3亚基I、50S核糖体蛋白L1[ 36], 叶绿体翻译延长因子EF-Tu[ 30, 31]和线粒体延长因子Tu[ 28]; 在蛋白折叠中涉及的蛋白dnak-类型的分子伴侣热休克蛋白(HSP) 70[ 33], 线粒体HSP 60[ 31]和新生多肽复合物结构域蛋白[ 46]; 与蛋白水解有关的蛋白包括20S蛋白酶体α-亚基[ 46]、α-类型5、α-类型6[ 42]和α-类型7[ 36], 26S蛋白酶体调节亚基4同源物和6A同源物[ 41, 42]等在种子萌发过程中增加积累。
信号转导相关蛋白, 14-3-3蛋白和14-3-3家族蛋白[ 39], 类14-3-3蛋白[ 29], 核苷二磷酸激酶和激酶I[ 33, 41, 42], 蛋白激酶APK18和假定的蛋白激酶[ 35, 45], 依赖-Ca2+蛋白激酶、腺苷酸激酶和假定的丝氨酸/苏氨酸蛋白激酶[ 42]也在种子萌发过程中增加积累。尽管这些蛋白在种子萌发过程中被增加积累, 但它们在种子萌发完成中的作用仍然不清楚。
液泡ATPase β-亚基、依赖电压的阴离子通道[ 28, 41, 42]和P-类型H+-ATPase[ 39]在种子萌发过程中增加积累, 这些蛋白与运输有关, 可能通过改变细胞的水势, 从而促进细胞伸长。与细胞生长和结构有关的蛋白, 例如肌动蛋白1和7[ 28, 30, 31, 42], 肌动蛋白解聚因子[ 52]和类肌动蛋白解聚因子蛋白[ 36], α-2,4、α-3,5和β-2微管蛋白[ 30, 31, 46], 微管蛋白α-1链和β-1链[ 28], 以及β-伸展蛋白(β-expansin)[ 41, 42]在种子萌发过程中增加积累, 表明这些蛋白与种子的萌发直接相关。拟南芥野生型种子中的α-2,4微管蛋白的丰度较低, 随着种子萌发其丰度明显增加; 多效唑(paclobutrazol)抑制种子萌发, 多效唑处理的种子中α-2,4微管蛋白的丰度类似于干燥种子。拟南芥GA缺陷型突变体种子( ga1 mutant seed)中的α-2,4微管蛋白的丰度类似于野生型干燥种子, 但不能萌发; 当在萌发介质中添加GA4+7时, 种子的萌发率达100%, α-2,4微管蛋白的丰度显著增加, 远远高于完全萌发的野生型种子[ 31]。水引发(hydropriming)和渗透引发(osmopriming)促进拟南芥种子的萌发速率和β-2微管蛋白的增加积累[ 30]。乙烯生物合成的关键酶1-氨基环丙烷-1-羧酸氧化酶(1-aminocyclopropane-1-carboxylate oxidase, ACO)在种子萌发过程中也增加积累[ 36, 39], 已经证明乙烯能促进种子萌发和拮抗ABA的作用[ 47]。
已经提出萌发过程本身就是一种“胁迫”, 细胞防御与救援蛋白的积累可能是种子萌发过程中活化的自我防御机制的一部分[ 42], 在种子萌发中起间接的(保护)作用。此外, 在种子萌发过程中, 增加积累一些贮藏蛋白; 根据这些蛋白的实验分子量小于理论分子量, 推测它们可能是贮藏蛋白的降解产物。应该注意的是, 在种子萌发过程中, 还有不同类型的蛋白下调, 其中一部分蛋白可能与种子萌发有关, 这里因篇幅的限制, 就不讨论了。
在干燥种子和萌发的种子中, 有许多蛋白受到翻译后修饰[ 45, 48], 这些修饰通过影响蛋白质的功能包括定位、复合物的形成、稳定性和活性等调节种子的萌发。
2.3.1 氧化还原信号 萌发伴随着种子中蛋白质氧化还原状态的广泛变化。在禾谷类作物中, 干燥种子的淀粉胚乳和胚中的蛋白质主要以氧化状态(S-S)存在, 在吸胀后被转化成为还原或者巯基(-SH)状态[ 49]。硫氧还蛋白(thioredoxin, Trx)是一种调节二硫键的蛋白, 在氧化-还原作用的转换中起重要作用。当烟酰胺腺嘌呤二核苷酸磷酸(NADPH)存在、Trx被酶促还原时, Trx作为一种萌发早期的信号起作用, 通过( a)还原贮藏蛋白, 增加它们的可溶性和对蛋白水解的敏感性; ( b)减少和失活抑制专一淀粉酶和蛋白酶的二硫健蛋白, 促进贮藏淀粉和蛋白的降解; 以及( c)还原性地活化在种子萌发中具有功能的酶[ 49], 促进贮藏物的动员。在模式豆科植物苜蓿种子的萌发过程中已经观察到类似的结果, 表明Trx像在单子叶植物中所描述的那样在双子叶植物种子萌发过程中起作用[ 50]。这些研究提出在成熟干燥种子中蛋白质组的氧化形式引起代谢的静止; 而种子萌发过程中在Trx存在时通过蛋白质二硫键的还原, 这些氧化形式的蛋白质被还原[ 49]。
另一种蛋白质的氧化修饰产生于ROS引起的蛋白质羰基化作用(protein carbonylation)。在拟南芥种子的吸胀过程中, 不同的羰基化蛋白积累[ 32]。这个过程的靶子是专一的代谢酶、翻译因子和一些分子伴侣。尽管在各种模式系统中常常认为羰基化蛋白的积累是在老化的背景下产生; 显然在种子中不是这样, 因为含有大量羰基化蛋白的拟南芥种子能够迅速地萌发和产生有活力的幼苗。似乎观察到的蛋白质羰基化模式的专一变化对于抵消和/或者利用由萌发种子代谢活性恢复所产生的ROS是需要的[ 51]。观察到的高丰度种子贮藏蛋白的羰基化作用可能反映了它们在清除种子萌发过程中产生的活性氧的作用[ 32]。此外, 已经提出这种羰基化作用可能通过降低蛋白结构的稳定性和促进蛋白水解来增加幼苗建成过程中种子贮藏蛋白的动员[ 32]。
2.3.2 磷酸化/去磷酸化 在细胞信号中磷酸化和去磷酸化代表一种普遍的调节机制。已经证明一组蛋白磷酸酶和蛋白激酶通过ABA信号的调节控制种子萌发[ 26, 52]。同样, 在DNA修复[ 45]和蛋白质翻译[ 53]中涉及的酶的磷酸化对于种子萌发的分子控制也是非常重要的。
2.3.3 一氧化氮(NO)介导的翻译后修饰 NO是生物系统中的一种主要的和多功能的中介物。然而, 尽管已经证明NO调节种子的代谢活性, 但它在种子萌发控制中的作用方式仍然不清楚。已经报道, NO能与金属蛋白的过渡金属结合(金属的亚硝基化)或者引起半胱氨酸的 S-亚硝基化作用( S-nitrosylation)或者酪氨酸的硝化作用(tyrosine nitration)[ 48, 54]。在吸胀1 h的拟南芥种子中, 已观察到NO的短暂爆发[ 55]。此外, 在吸胀24 h的高粱胚轴中已观察到一些硝化蛋白增加[ 56]。蛋白硝化作用(protein nitration)是一种硝化应激(nitrosative stress)的生物学标记, 可能参与植物中蛋白质的转换或者信号转导[ 57, 58]。有趣的是, 钼辅因子硫化酶(molybdenum cofactor sulfurase, ABA3, ABA合成的最后步骤涉及的一种酶)的酪氨酸硝化作用已经在拟南芥中报道[ 59]。在种子吸胀过程中, 由这种机制引起的ABA合成的失活可能有助于种子的萌发。此外, 增加的证据表明蛋白质的 S-亚硝基化作用可能调节种子萌发过程中的代谢和能量过程[ 48]。
除了上述翻译后修饰外, 已经证明蛋白质生物素化(biotinylation)、糖基化(glycosylation)、泛素化(ubitinylation)、法尼基化(farnesylation)和乙酰化(acetylation)也对种子萌发起调节作用[ 48]。
脱落酸是种子休眠的正调节剂, 是萌发的负调节剂, 在种子休眠和萌发中起主要作用[ 47, 60, 61]。许多ABA缺陷型的物种表现出萌发能力增加, 有时产生胎萌的种子; 而过量积累ABA的突变体和转基因系表现出加强休眠[ 62, 63]。这些基因型已经证明了ABA在种子发育过程中的主要功能是抑制提前萌发和诱导初生休眠。同样, 利用ABA生物合成抑制剂的实验表明, 在吸胀种子中ABA的重新合成对于种子休眠的维持是必需的[ 64]。
细胞内的ABA含量被生物合成和降解代谢之间的平衡所调节[ 65]。9-顺式-环氧类胡萝卜素二加氧酶(9-cis-epoxycarotenoid dioxygenases, NCED)催化9-顺式-环氧类胡萝卜素、9-顺式-新黄质和9’-顺式堇菜黄质的裂解, 这是ABA生物合成的主要调节步骤。玉米黄质环氧酶(zeaxanthin epoxidase, ZEP), 一种叶黄素环化酶, 也被认为在ABA生物合成过程中具有调节功能。ABA被羟基化作用或者与糖结合失去活性。P450单加氧酶的CYP707A家族成员编码ABA 8’-羟化酶(8’-hydroxylase), 这是ABA 8’-羟化作用途径中的关键步骤, 在ABA含量的控制中起调节作用[ 47, 60]。到目前为止研究过的所有物种都以多基因家族的形式编码NCED和CYP707A, 这些成员的不同结合有助于组织和环境的专一调节。
在发育过程中积累和在干燥种子中存在的ABA随着种子吸胀下降。这种下降在休眠(Cvi)和非休眠(Col)的拟南芥种子中发生, 主要取决于CYP707A2的活性[ 62, 64, 66, 67, 68]。在Col和Cvi拟南芥种子中, CYP707A2基因在吸胀开始后2~3 h被诱导[ 68], 导致ABA含量的迅速下降, 暗示与CYP707A2蛋白的重新合成关系密切[ 67]。这种早期诱导被几种因子, 例如硝酸盐[ 69]、NO[ 67]和后熟[ 81]所调节。休眠的Cvi种子和热抑制的Col种子在吸胀初期表现出ABA含量下降, 然后增加[ 64, 70]。因此, ABA含量的降低可能是种子萌发的一个前提条件。
拟南芥分子遗传学的最近进展揭示了核心的ABA信号途径。蛋白磷酸酶2C (protein phosphatase 2C, PP2C)基因家族的成员, 包括ABA不敏感I ( ABA-INSENSITIVE1, ABI1)和 ABI2是中心调控因子(图3-A), 这些基因功能的缺失突变引起ABA的过敏性反应, 暗示它们是ABA信号的负调控因子[ 71]。隐性PP2C突变体的比较分析表明, AtPP2CA/AHG3是吸胀种子中的重要执行者[ 72]。蔗糖非发酵-1 (sucrose non-fermenting-1, SNF1)相关的蛋白激酶亚家族2 (SNF1-related protein kinase subfamily 2, SnRK2)的成员是一种ABA信号的关键正调控因子, snrk2.2 snrk2.3 snrk2.6三突变体在萌发过程中以及在高湿度下的胎萌中表现出一种强烈的ABA不敏感性[ 73, 74]。SnRK2能活化ABA反应元件(ABA-responsive element, ABRE)结合的碱性亮氨酸拉链(basic leucine zipper, bZIP)转录因子, 包括ABI5[ 74]。当SnRK2从PP2C的抑制中去抑制时, 成为活性形式[ 75, 76]。ABA的受体蛋白有5种不同的形式[ 77, 78, 79, 80]。在这些受体中, 起始蛋白(START protein)的PYR1/PYL/RCAR家族以依赖于ABA的方式通过调节PP2C的活性在种子ABA响应中起控制作用[ 78, 80]。在拟南芥基因组中, 家族的14个成员(PYR1, PYL1-13)被编码, pyr1 prl1 prl2 p rl4四重突变体表现出强烈的ABA不敏感萌发[ 80]。
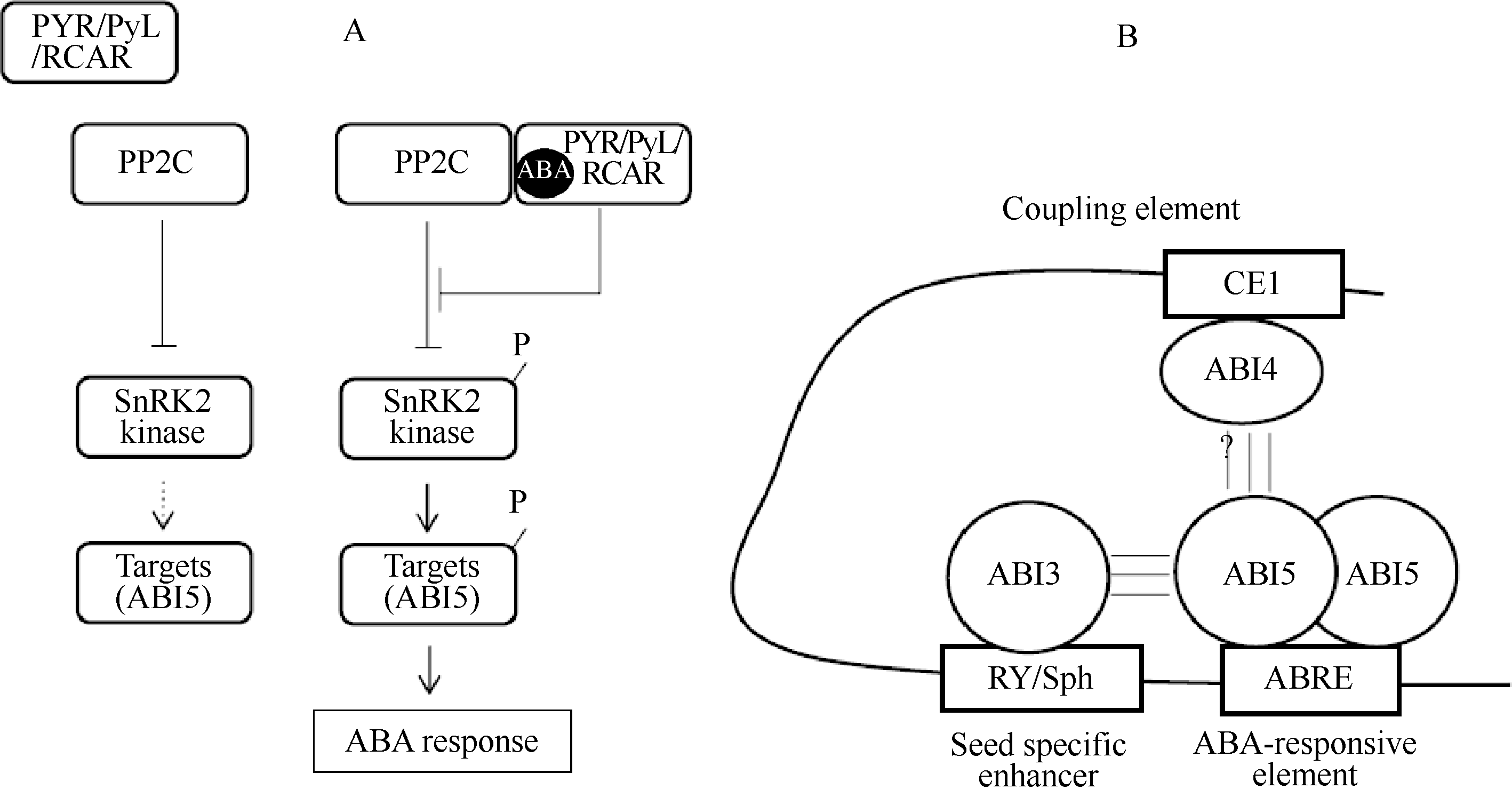 | 图3 ABA信号途径Fig. 3 The pathway for ABA signalling |
在种子中, 转录作用是ABA响应中的重要步骤(图3-B)。遗传分析表明ABI3 (B3类型)、ABI4 (AP2类型)和ABI5 (bZIP类型)是授予种子ABA响应的关键转录因子[ 81]。在植物界, 这些转录因子是保守的, 拟南芥中相应的直系同源基因也存在于单子叶植物中。ABI5/TRAB1专一地与典型的ABRE结合, 这是触发ABA介导的转录作用的顺式元件。在吸胀的拟南芥种子中, ABI5蛋白被ABA调节, 是一种对ABA响应的指示物[ 82]。ABI3/VP1与RY/Sph (一个种子专一的启动子)重复结合, 而ABI4/ZmABI4与耦合元件1 (coupling element 1, CE1) 结合, 这些因子与ABRE一道起作用以触发ABA介导的转录作用[ 60]。
在种子萌发过程中, GA能促进种子萌发和拮抗ABA的抑制作用[ 47]。具有生物活性的GA4存在于干燥种子和后熟种子中, 在种子萌发后期GA4含量进一步增加[ 83]。GA20氧化酶(GA20ox)和GA3氧化酶(GA3ox)是GA生物合成的关键调节酶, 而GA2氧化酶催化GA的降解[ 84]。Ogawa等[ 83]证明, 在种子萌发过程中, GA的生物合成定位于胚根、下胚轴和珠孔端胚乳。单子叶和双子叶植物的GA生物合成酶突变体是对GA敏感的, 表现出生长和发育的缺陷, 这些缺陷能被外源GA恢复。水稻和大麦的GA敏感突变体表现出矮小、不育以及种子萌发过程中不能通过α-淀粉酶的诱导动员贮藏物[ 85]。编码GA降解代谢酶 GA 2 ox的基因的过量表达增加GA的转变, 导致减少小麦种子的萌发和α-淀粉酶的诱导[ 86]。
GA信号由一种可溶性受体蛋白GA不敏感的矮秆蛋白1 (GA-INSENSITIVE DWARF1, GID1)感受。感受GA的机制是保守的, 在拟南芥和水稻中是一致的[ 84]。通过图位克隆(map-based cloning)技术, 已经在水稻GA不敏感的突变体中鉴定了 GID1基因[ 87], 在大麦中鉴定了 GID1的同系物 GSE1[ 88], 以及在拟南芥中鉴定了同系物 GID1a、 GID1b和 GID1c[ 89, 90]。GA受体的突变产生类似于严重的GA生物合成突变的表型, 但这些表型不能被外源GA恢复。GID1蛋白主要定位于细胞核, 但也有定位细胞质的报道[ 90]。
DELLA基因被定义为GA信号的阻遏物, 因为 DELLA基因的功能获得突变(gain-of-function mutation)引起减少的GA信号(例如矮小), 而功能丧失突变(loss of function mutation)导致增加的GA信号(例如苗高或者细长的表型)。GID1通过直接的蛋白质-蛋白质相互作用增加DELLA的阻遏反应。所有的DELLA阻遏物含有一个保守的氨基酸顺序的N-末端DELLA调节区域(Asp-Glu-Leu-Leu-Ala)和一个C-末端GRAS (例如GAI、RGA和SCARECROW)功能区域[ 84]。大麦的 DELLA基因被称为 SLENDER1 ( SLN1), 水稻的 DELLA基因被称为 SLENDER RICE1 ( SLR1)。拟南芥具有5个 DELLA基因, 其中 REPRESSOR OF GA1-3 ( RGA)、 GA-INSENSITIVE( GAI)和 RGA-LIKE ( RGL1)的突变能够救援(rescue)植株的高度, RGA、RGL2和 RGL1的突变能够救援开花, 而 RGL2、 RGA、 GAI和 RGL3的突变能够救援种子的萌发; DELLA RGL2显著地影响种子萌发, 而DELLA RGA显著地影响植株高度[ 84]。
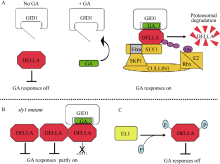
GA能通过泛素-蛋白酶体途径(ubiquitin- proteasome pathway)目标水解DELLA蛋白, 从而降低DELLA对GA反应的阻遏作用(图4-A)。最初, 这个模型的根据是GA缺乏所引起的矮小表型能被外源GA处理救援, 这种救援与DELLA蛋白的消失有关[ 91]。SLY1/GID2是催化DELLA蛋白多聚泛素化的SCF(SKP1、CULLIN和F-BOX) E3泛素连接酶的F-box亚基[ 92, 93]。SCF复合物的F-box亚基通过一个C-末端的蛋白质-蛋白质相互作用区域授予底物特异性, 以及通过N-末端的F-box区域与SCF复合物的其余部分相互作用[ 94]。用GA处理GA生物合成突变体 ga1-3, 种子在5 h内或者幼苗在10 min内, 通过泛素化作用导致DELLA蛋白的破坏[ 95, 96]; 这种破坏能够被26S蛋白酶体的抑制剂阻断[ 97]。因此, GA促进的DELLA蛋白的多聚泛素化作用通过26S蛋白酶体使其成为靶子被降解。
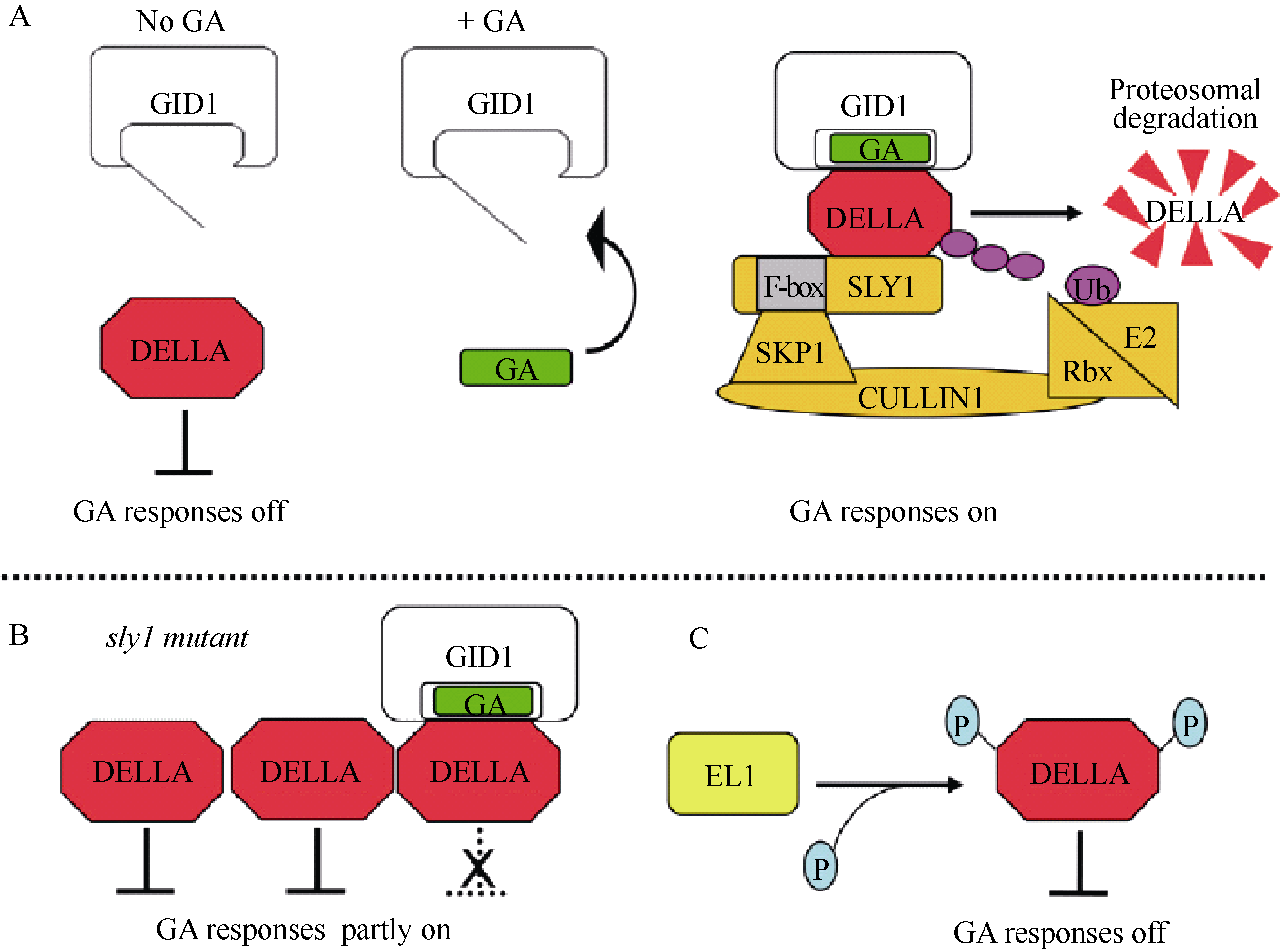 | 图4 依赖和不依赖蛋白水解的GA信号模型Fig. 4 Proteolysis-dependent and -independent GA signaling models |
GID1-GA-DELLA复合物的形成触发DELLA和F-box蛋白SLY1/GID2之间的蛋白质-蛋白质相互作用。酵母三-杂交分析(yeast three-hybrid analysis)表明, GID1-GA-DELLA复合物的形成促进GID2与DELLA的结合[ 98]。GA促进SLY1与GID1的共免疫沉淀(coimmunoprecipitation), 暗示GID1-DELLA复合物的形成也促进植物中GID1-DELLA-SLY1的相互作用[ 92]。在GA生物合成突变体 gid1和 sly1/gid2中DELLA蛋白的高水平积累表明GA、GID1受体和SCF复合物的组分对于DELLA蛋白通过泛素-蛋白酶体途径水解是需要的[ 97]。此外, 在无细胞系统中, GA、GID1、SCFSLY1和泛素足够使拟南芥DELLA蛋白发生多聚泛素化作用。在拟南芥和水稻中有一个SLY1/GID2的同系物称为SNEEZY (SNE)或者SLY2[ 99]。SNE可能在GA信号中起作用, 而SLY1是DELLA蛋白积累的主要调控因子, 已知在 sly1 sne双突变体中 SN E突变具有小的加性效应[ 92, 100]。
如果所有GA信号通过DELLA蛋白水解发生, 那么, DELLA蛋白积累的水平应该总是与严格的GA不敏感表型相关。在 sly1和 gid2 F-box突变体中的情况并不是这样, 它积累多得多的DELLA蛋白, 但表现出比GA生物合成或者GA受体无效的突变体要较少严格的GA不敏感表型[ 90, 101]。似乎DELLA能被一种非蛋白水解机制失活, 这样不是所有的在 sly1/ gid2突变体中积累的DELLA蛋白在功能上作为GA信号的阻遏物[ 95, 101, 102]。根据下面的证据提出了GID1-GA-DELLA复合物的形成单独阻断DELLA抑制的模型: (1) sly1/ gid2表型被 GID1基因的过量表达救援, 而没有DELLA蛋白水平的降低; (2) sly1/ gid2中的GA信号需要GID1-GA-DELLA复合物形成所需要的所有元件, 包括GA激素的合成、 GID1基因、DELLA-GID1蛋白相互作用需要的DELLA区域。根据这种情况, 似乎在 sly1突变体中GID1通过与DELLA蛋白结合而不是通过触发DELLA蛋白水解引起GA信号(图4-B)。不像拟南芥GID1a和GID1c那样, GID1b蛋白与GA的结合具有较高的亲和性, 在GA缺乏时表现出能与DELLA相互作用。这就解释了为什么 GID1b的过量表达能更好地援救GA不敏感的 sly1突变体表型的原因[ 102, 103]。在GA缺乏时, 这种GA-不依赖的 GID1b-DELLA结合可能提供了一种低水平的GA信号。拟南芥GID1b具有低的表达水平, 暗示GID1b积累的控制对于调节基本的GA反应可能是必需的[ 90, 104]。
已有的证据表明, DELLA的活性可能被磷酸化和 O-连接的N-乙酰葡糖胺( O-linked N-acetylglu-cosamine, O-GlcNAc)修饰影响。研究表明DELLA被磷酸化, 但这种修饰在控制DELLA活性或者蛋白降解中的精确作用是不清楚的。DELLA的磷酸化最初在水稻和拟南芥中观察到[ 94, 105]。最近的证据表明, 由水稻 EARLY FLOWERING1 ( EL1)编码的丝氨酸/苏氨酸蛋白酪蛋白激酶I (Ser/Thr protein casein kinase I)通过直接的蛋白磷酸化作用调节DELLA SLR1[ 106]。 EL1作为GA信号的负调节因子和DELLA的正调节因子起作用, 因为丧失 EL1功能导致与早期开花有关的GA敏感性增加以及引起种子萌发过程中α-淀粉酶的增加表达。 El1表型与GA处理后DELLA SLR1蛋白降解速率的轻微增加有关; 与磷酸酶抑制剂的实验一致, 磷酸化的DELLA更稳定。两方面的证据建议, EL1通过磷酸化调节DELLA。首先, EL1蛋白在体外磷酸化DELLA SLR1。其次, 在DELLA SLR1 (S196A和S510A)中Ser到Ala的突变能阻断EL1-依赖的磷酸化作用, 防止基因表达的DELLA活化。此外, Ser到Asp的突变(S196D和S510D)导致DELLA活化的转录物增加积累。这就提出DELLA可能通过这2个位点的磷酸化被调节, 一个位点在DELLA调节区域内, 另一个位点在GRAS功能区域内[ 84]。
SPINDLY ( SPY)编码一种 O-GlcNAc转移酶, 在拟南芥、水稻和小麦中负调节GA信号[ 107, 108, 109]。在拟南芥中, spy抑制功能获得的DELLA突变 rga-Δ17。这种抑制与DELLA的消失无关, 而与明显的DELLA磷酸化有关[ 110]。 SPY RNA干扰也抑制水稻 gid2 F-box突变体, 而没有DELLA的蛋白水解, 但引起DELLA SLR1蛋白磷酸化的增加[ 108]。由于沉默 SPY也抑制水稻 gid1和GA生物合成突变体, 似乎在 spy中增加的GA信号既不需要DELLA的蛋白水解, 也不需要GID1-DELLA复合物的形成[ 84]。
在高等植物中, 乙烯通过Yang-cycle从Met合成[ 111]。乙烯生物合成的限速步骤分别由2个酶家族调节, ACC合酶(ACC synthase, ACS)催化AdoMet转换成为ACC; ACO催化ACC转变成为活性乙烯。ACO的积累和乙烯释放的增加与一些物种的种子萌发相联系[ 112, 113]。已经证明ACO转录物的水平被乙烯本身和其他植物激素调节[ 114]。在豌豆种子萌发过程中, 乙烯通过 Ps-ACO1转录物的正反馈调节促进乙烯的生物合成, 而 Ps-ACS1 mRNA的水平和ACC的总含量不被乙烯处理影响。乙烯对豌豆种子萌发的促进作用与β-1,3-葡聚糖酶有关, 此酶在胚轴的胚根中被专一地诱导表达, 但不在子叶中被诱导表达[ 115, 116]。乙烯的生物合成和响应性被定位在豌豆胚根的延长区和分化区。用乙烯产生试剂乙烯利处理钻果大蒜芥种子不显著地影响 S oACS7 mRNA的水平和ACC的总含量[ 117]。与这个结果一致, 由氨基乙氧基乙烯甘氨酸(aminoethoxyvinylglycine, AVG)抑制ACS对种子萌发没有明显的影响, 而Co2+抑制ACO减少钻果大蒜芥的最大萌发百分数[ 117]。这些结果表明, So A CO2的表达与钻果大蒜芥种子的萌发有关。
乙烯信号是通过它的受体发生的; 最初在拟南芥中分离到的受体突变体是乙烯三重反应1 ( et h ylene triple response 1, etr1), 一种定位于内质网膜的二聚体组氨酸激酶和乙烯信号的负调控因子[ 118]。在拟南芥中, 已知有ETR1、ETR2、ERS1、ERS2和EIN4五种乙烯受体。当与乙烯结合时, 受体失活, 相继使组成性三重反应1 (constitutive triple response1, CTR1)失活。CTR1是一种丝氨酸-苏氨酸蛋白激酶, 作为一种乙烯信号的负调控因子起作用。失活的CTR1引起连续的MAP-激酶级联(MAP-kinase cascade), 以及调节正调控因子EIN2和它定位在核中的转录因子, 例如EIN3、EIL1和EREBPs/ERFs, 从而活化乙烯反应基因的转录[ 119]。已经证明, 在乙烯作用抑制剂2,5-降冰片二烯(2,5-norbornadiene, NBD)的实验中, 已知的乙烯受体的信号对于许多物种的种子萌发是重要的, 例如十字花科的拟南芥和家独行菜、豆科的豌豆、茄科的烟草和许多其他物种[ 47, 119]。已经提出几种假说来解释乙烯在萌发种子中的作用机制, 家独行菜种子珠孔端胚乳弱化的生化定量强有力地表明乙烯信号和生物合成是这个过程所需要的[ 119]。通过诱导引起细胞壁松弛或者细胞分离的细胞壁重塑蛋白(cell wall remodeling proteins, CWRP)和/或者ROS的表达, 表明乙烯促进珠孔端胚乳的弱化[ 61, 120]。已经提出, 在胚轴中乙烯通过促进径向细胞膨胀或者通过降低种子的基础水势(base water potential)起作用[ 47, 118]。
种子萌发的控制是一个非常复杂的过程, 它需要不同激素之间的协同作用和相互作用[ 47]。已知种子的休眠状态被ABA诱导和维持, 而被GA解除。在更宽的植物休眠的视野中, 这种ABA-GA的拮抗作用也是明显的, 它控制块茎和休眠芽的萌发[ 121, 122]。ABA和它相关的转录因子例如ABI3/VP1被描述为古老的休眠调控因子, 因为在系统发生上它们是普遍的, 因此, 在进化上它们是古老的[ 61, 123, 124]。与ABA相反, GA负调节休眠, 因为它解除种皮介导的种子休眠和促进种子萌发[ 47, 123]。ABA和GA代谢在种子萌发过程中相互联系, 例如在 aba2-2突变体中GA的生物合成增加, ABA抑制种子萌发过程中GA的生物合成[ 125]。此外, 与野生型比较, 在 ga1突变体中ABA合成更多, 降解更少; 外源添加GA后, 相反的结果被观察到[ 81]。我们的结果也表明ABA显著地抑制莴苣种子的萌发, 但这种抑制作用能被GA3有效地降低[ 126]。
乙烯促进种子萌发, 以及拮抗ABA对种子萌发的抑制作用[ 47, 119]。乙烯不敏感 etr1突变体种子表现出形状改变、休眠增加、萌发率低以及它们的萌发是ABA过敏性的[ 112]。在 etr1种子中, 这种现象至少部分是由于较高的ABA含量引起的。同样, 具有降低乙烯敏感性的 ein2突变体种子表现出增加的ABA敏感性[ 112]。已经证明, 拟南芥 ctr1功能丧失突变体种子萌发过程中具有减少的ABA敏感性, 以及在ACC介导的胚乳破裂和ABA抑制的恢复中是受损伤的[ 119]。
已经提出, 在解除休眠和促进萌发中存在一种GA-乙烯的增效作用[ 118]。在打破欧洲水青冈种子休眠过程中, GA20氧化酶( FsGA 20 ox1)的表达被连接到这两种激素之间的交叉作用[ 127]。在钻果大蒜芥中, 当 SoACO2的表达被种子后熟抑制时, GA3氧化酶( SoGA 3 ox)的表达被促进[ 128]。在GA缺乏的拟南芥 ga1-3突变体中, GA处理导致ACO表达的增加, 这就提出GA活化乙烯的生物合成和/或者反应。此外, 在 ga1-3突变体中, 乙烯处理能够代替GA缺乏和诱导在光下萌发。与这些结果相反, 乙烯不能代替GA处理诱导GA缺乏的 gib-1番茄突变体种子的萌发[ 119]。
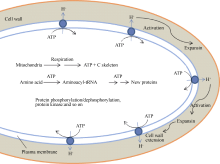
种子萌发是一个复杂的过程, 在这个过程中静止的干燥种子迅速地恢复代谢活性, 完成胚伸出周围结构的细胞事件, 以及为随后的幼苗生长做准备[ 1, 2, 3, 6]。在拟南芥、油菜、苜蓿、玉米等种子萌发之前, 胚轴细胞的伸长是显著的, 但未观察到明显的细胞分裂[ 129, 130, 131, 132]。在萌发过程中, 种子大小和形状的所有变化都是由细胞膨胀引起的。膨胀的植物细胞通过改变壁的主要组分纤维素微纤丝和果胶/半纤维素调整细胞壁的延展性。细胞壁的松弛允许水分渗入, 从而驱动细胞膨胀和产生细胞膨压[ 133]。这就提出在萌发过程中胚的伸长可能主要是依赖于细胞壁延展性的增加。Bethke等[ 134]在拟南芥种子水分吸收的第二阶段早期观察到胚和胚乳细胞具有许多小液泡。已经观察到, 拟南芥种子的萌发不是由于胚根本身的延长引起的, 而是由于下胚轴以及下胚轴和胚根之间的过渡区的少量细胞伸长引起的[ 132]。因此, 根据种子萌发过程中下胚轴-胚根细胞的伸长、代谢活性的迅速增加以及由差异蛋白质组分析所构建的关键事件, 我们提出了“种子萌发的能量刺激假说”(图5)。
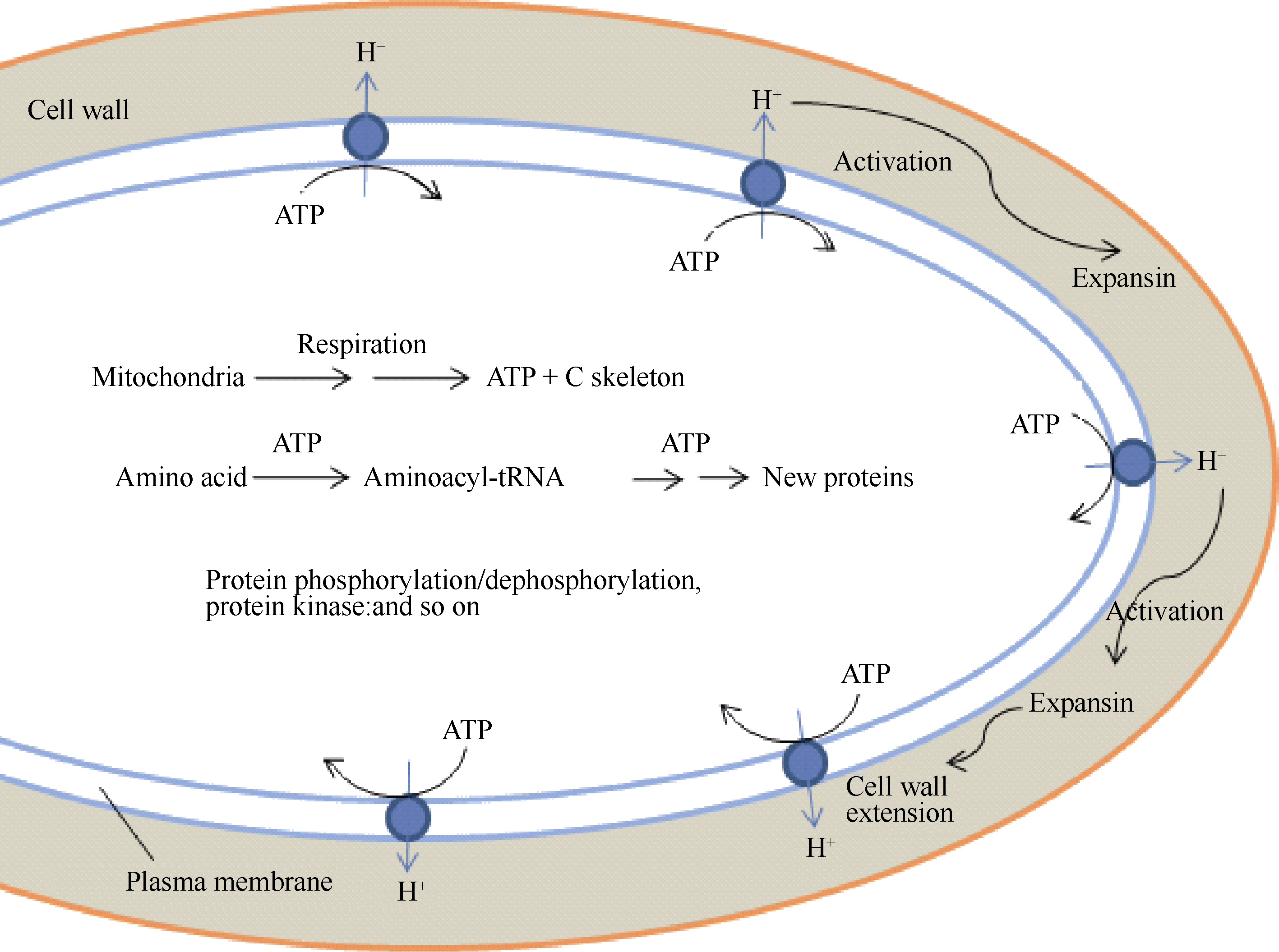 | 图5 种子萌发的能量刺激假说Fig. 5 Energy stimulating hypothesis for seed germination |
种子萌发的能量刺激假说认为在干燥种子吸胀初期, 预存线粒体呼吸作用(能量产生)的迅速增加在种子萌发中起原初作用, 但不足以使种子完成萌发, 需要一系列事件的协同作用, 才能引起细胞壁松弛、细胞伸长和完成萌发(图5)。胚轴细胞质膜具有H+-ATPase, 能水解呼吸作用产生的ATP, 同时将H+泵入细胞壁, 从而活化伸展蛋白(expansin), 使细胞壁松弛, 细胞吸水和伸长(图5)。已经提出伸展蛋白是促进细胞壁膨胀的细胞外蛋白, 可能通过破坏半纤维素细胞壁组分和纤维素微纤丝之间的氢键起作用[ 135]。 LeEXP4mRNA专一地定位于珠孔端胚乳帽区域, 表明这种蛋白可能有助于胚根伸出需要的组织弱化。GA缺乏的( gib-1)突变体种子只有在外源GA存在时才能萌发, 在吸胀12 h内GA诱导 LeEXP4的表达[ 135, 136]。此外, 氨基酸的活化, 蛋白质的合成、降解和磷酸化修饰也需要ATP; 呼吸作用的中间产物能为其他生物大分子的合成提供碳骨架。值得注意的是种子萌发的能量刺激假说是根据近年来的研究进展结合作者实验室的研究结果提出来的, 还需要进一步用实验来证实。可以认为, 此假说将为解释种子萌发的机制和减少穗萌提供新的研究思想。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|