

 1V (1A) and CS
1V (1A) and CS{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
簇毛麦新型HMW-GS的序列分析及加工品质效应鉴定
[杨华1 , 高翔1, 2, *  , 陈其皎
, 陈其皎1 , 赵万春1, * , 董剑1, 2 , 李晓燕1 ]
, 陈其皎, 董剑|
|


利用设计的HMW-GS特异引物, 从簇毛麦TA10220中克隆到6条HMW-GS基因序列(1.0~1.7 kb, GenBank登录号为KF887414~KF887419), 比普通小麦HMW-GS基因序列小, 其中KF887414~KF887417为能正常表达的基因, KF887418和KF887419是编码区出现终止信号的假基因。综合推导氨基酸的序列分析以及基于编码蛋白全序列、N-端和C-端域的进化树分析, 认为KF887414属于y型HMW-GS, KF887415~KF887419的氨基酸序列同时具有x和y型的结构特征。将具有完整编码区的4个基因在宿主菌
小麦高分子量谷蛋白亚基(HMW-GS)和低分子量谷蛋白亚基(LMW-GS)通过分子内和分子间二硫键形成聚合蛋白, 从而影响面粉的加工品质[ 1]。HMW-GS影响面团的弹性和面包的加工品质[ 2, 3]。编码HMW-GS (60~120 kD)的基因位于第1部分同源群染色体1A、1B和1D长臂上[ 4, 5, 6, 7, 8, 9], 每个位点由2个紧密连锁的基因组成, 其中较大的基因编码x型HMW-GS的合成, 较小的基因编码y型HMW-GS的合成[ 9, 10]。HMW-GS具有高度保守的5’和3’端序列, 编码区内部不存在内含子[ 11, 12]。HMW-GS典型的结构由4部分组成, 分别是信号肽(蛋白质加工过程被删除)、高度保守的N-端和C-端以及中间重复区[ 2, 3]。x和y型HMW-GS主要区别在于半胱氨酸残基的个数和位置, 中间重复区重复单元的类型, 其中六肽以PGQGQQ、九肽以GYYPTS/PLQQ的保守形式出现在所有x、y型亚基中, 而三肽以GQQ的保守形式只存在在x型亚基中[ 2, 3]; 多数的半胱氨酸残基存在于N-端保守区, y型的有5个, 并且第3和第4个相邻, x型的有3个; C-端都只有1个; 但是1By9、1Dy10和1Dy12近C-端的中间重复区多了1个半胱氨酸残基, 而1Dx5近N-端的中间重复区多了1个半胱氨酸残基[ 3]。
簇毛麦( Dasypyrum villosum, 2 n = 14, VV)属于禾本科小麦亚族簇毛麦属, 为一年生二倍体异花授粉草本植物, 具有抗多种病害、品质优良、抗旱耐寒、密穗、多花及分蘖能力强等特性, 还带有提高籽粒蛋白质含量、赖氨酸含量和面筋强度的基因, 是普通小麦遗传改良优异的三级基因源[ 13, 14]。簇毛麦1V染色体对小麦加工品质具有正效应, 可以显著提高面筋的强度, 是位于1V染色体上 Glu-V1、 Gli-V1和 Glu-V13位点的作用[ 15, 16, 17, 18]。其中1V染色体上有: 编码HMW-GS的 Glu-V1位点, 编码贫硫(ω型)和富硫(γ型)醇溶蛋白基因 Gli-V1位点和编码低分子谷蛋白 Glu-V3位点[ 15, 16, 17, 18, 19, 20]。
利用同源克隆技术, 一些研究者从簇毛麦中分离到HMW-GS编码基因及其启动子序列, 并发现了没有功能的假基因[ 20, 21, 22, 23, 24]。迄今, 尚缺乏对该基因家族成员多样性、序列特征、以及加工品质特征方面的深入报道。对簇毛麦HMW-GS基因序列和推导的氨基酸序列分析, 可为进一步了解簇毛麦HMW-GS与普通小麦HMW-GS的关系提供依据, 而且原核体外表达和功能鉴定有助于验证簇毛麦HMW-GS对小麦籽粒品质和加工品质的影响。
参考段淑娥等[ 25]和Verbruggen等[ 26]的方法, 从单粒种子中提取簇毛麦TA10220 (美国堪萨斯州大学小麦遗传资源研究中心提供) HMW-GS, 然后进行SDS-PAGE分析[ 25, 26]。
以室内培养2周的簇毛麦TA10220幼嫩叶片为材料, 采用微量CTAB法提取基因组DNA[ 27]。
根据GenBank公布的HMW-GS基因编码区的保守序列设计1对简并引物(hmw-F: 5'-TCATCACC CACAACACCGA-3'; hmw-R: 5'-TCACTGGCTRGC CGACAAT-3'), 以簇毛麦TA10220的基因组DNA为模板扩增目的基因。反应体系25 μL, 包含5× PrimeSTAR buffer (Mg2+ plus) 5 μL, dNTP Mixture (各2.5 mmol L-1) 2 μL, 引物各1 μL, DNA模板1 μL, PrimeSTAR HS DNA Polymerase (2.5 U μL-1, TaKaRa) 0.2 μL。PCR反应条件为94℃ 2 min; 98℃ 10 s, 68℃ 30 s, 72℃ 2 min, 共30个循环; 最后72℃延伸10 min。
PCR产物经琼脂糖凝胶电泳检测, 利用DNA凝胶回收试剂盒(天根生化科技北京有限公司)回收目的条带, 对回收的产物先进行末端加A反应, 其反应体系10 μL, 含回收产物4 μL, 2 mmol L-1dATP 0.8 μL, 10×PCR buffer 1 μL, Taq DNA polymerase 0.3 μL; 然后与克隆载体pEASY-T1 (北京全式金生物技术有限公司)连接, 转入克隆菌株 E. coli Trans1-T1 (北京全式金生物技术有限公司)中, 使用T7/SP6通用引物对白色菌落进行PCR扩增, 筛选阳性克隆, 由南京金斯瑞生物科技有限公司测序。使用DNAMAN、MEGA5.0及NCBI和SignalP 4.1 Server网站上的在线工具对所得序列进行核苷酸及推导氨基酸序列分析。
根据测序结果设计2对表达引物(hmwExF1: 5'- GAAGGTGGGGCCTCTGC-3', hmwExR1: 5'-CTGG CTGACCGACAATGC-3'; hmwExF2: 5'-GAAGGCG AGGGCTCTGGG-3', hmwExR2: 5'-CTGGCTGGCC GACAATGC-3')。将表达引物扩增的目的基因与表达载体pEASY-E2 (北京全式金生物技术有限公司)连接, 转入克隆菌株 E. coli T rans1-T1中, 在LB固体培养基(含50 mg L-1 AMP)上筛选阳性克隆。用T7F/hmwExR或hmwExF/T7R引物筛选具有正确表达方向的重组子, 经测序确定序列的正确性。提取质粒, 转入表达菌株 E. coli Rosetta-gami B(DE3)(北京全式金生物技术有限公司)中。挑取单个阳性克隆至LB液体培养基(含50 mg L-1 AMP)中, 37℃过夜, 将培养物按1∶100接种至新鲜LB液体培养基(含50 mg L-1 AMP)中, 37℃, 200转 min-1培养至菌液OD600值为0.5; 取1 mL菌液作为阴性对照, 向剩余菌液中加入IPTG (1 mmol L-1), 37℃诱导6 h, 12 000 × g离心10 min收集菌体, 用SDS-PAGE (12%分离胶, 4%浓缩胶)分析蛋白表达结果。
处理后的蛋白样品经12% SDS-PAGE, 马上电转移至硝酸纤维素(NC)膜, 用新鲜配制的封闭液(5%脱脂奶粉), 在室温摇床封闭1 h, 弃封闭液; 以鼠抗His抗体为一抗(1∶10 000), 室温孵育1 h, 用TBS和TBST各洗膜2次, 每次15 min; HRP标记的羊抗鼠IgG为二抗(1∶10 000), 用TBS和TBST各洗膜2次, 每次15 min, 最后用DAB显色试剂盒进行化学发光显色。
收集诱导表达菌液, 于4℃ 12 000 × g离心 10 min, 弃上清液, 加入10 mL破碎缓冲液(pH 8.5, 50 mmol L-1 Tris-HCl, 2 mmol L-1EDTA, 100 mmol L-1 NaCl, 1 mg L-1溶菌酶)重悬菌体。冰浴45 min后超声破碎, 离心收集沉淀。沉淀经过预处理后过柱纯化, 平衡缓冲液为50 mmol L-1磷酸缓冲液(pH 7.4, 0.5 mol L-1 NaCl, 20 mmol L-1咪唑), 将洗脱缓冲液为50 mmol-1磷酸缓冲液(pH 7.4, 0.5 mol L-1 NaCl, 4 mol L-1脲, 500 mmol L-1咪唑), 收集的溶液透析24 h, 低温冷冻干燥保存待用。用SDS-PAGE (12%分离胶, 4%浓缩胶)分析蛋白纯化结果。
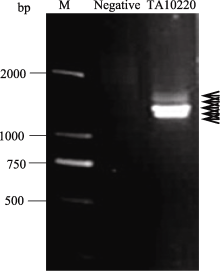
利用引物对hmw-F/hmw-R扩增簇毛麦TA10220基因组DNA, 获得1~2 kb的目标条带(图1)。回收该条带, 末端加A, 连接至pEASY-T1载体上, 转化到大肠杆菌Trans1-T1中, 经过菌落PCR扩增, 筛选出43个阳性克隆, 测序后通过序列比对, 得到6个非重复序列。用NCBI在线工具BLASTn对测序结果进行同源性分析, 表明测得6条序列与数据库中已经公布的一年生簇毛麦( D. villosum)、多年生簇毛麦( D. breviaristatum)、小麦及其亲缘植物的HMW-GS相似性很高。因此, 初步确定这6个序列为簇毛麦HMW-GS基因, 将序列提交GenBank (http://www.ncbi.nlm.nih.gov/sites/entrez), 登录号为KF887414至KF887419。
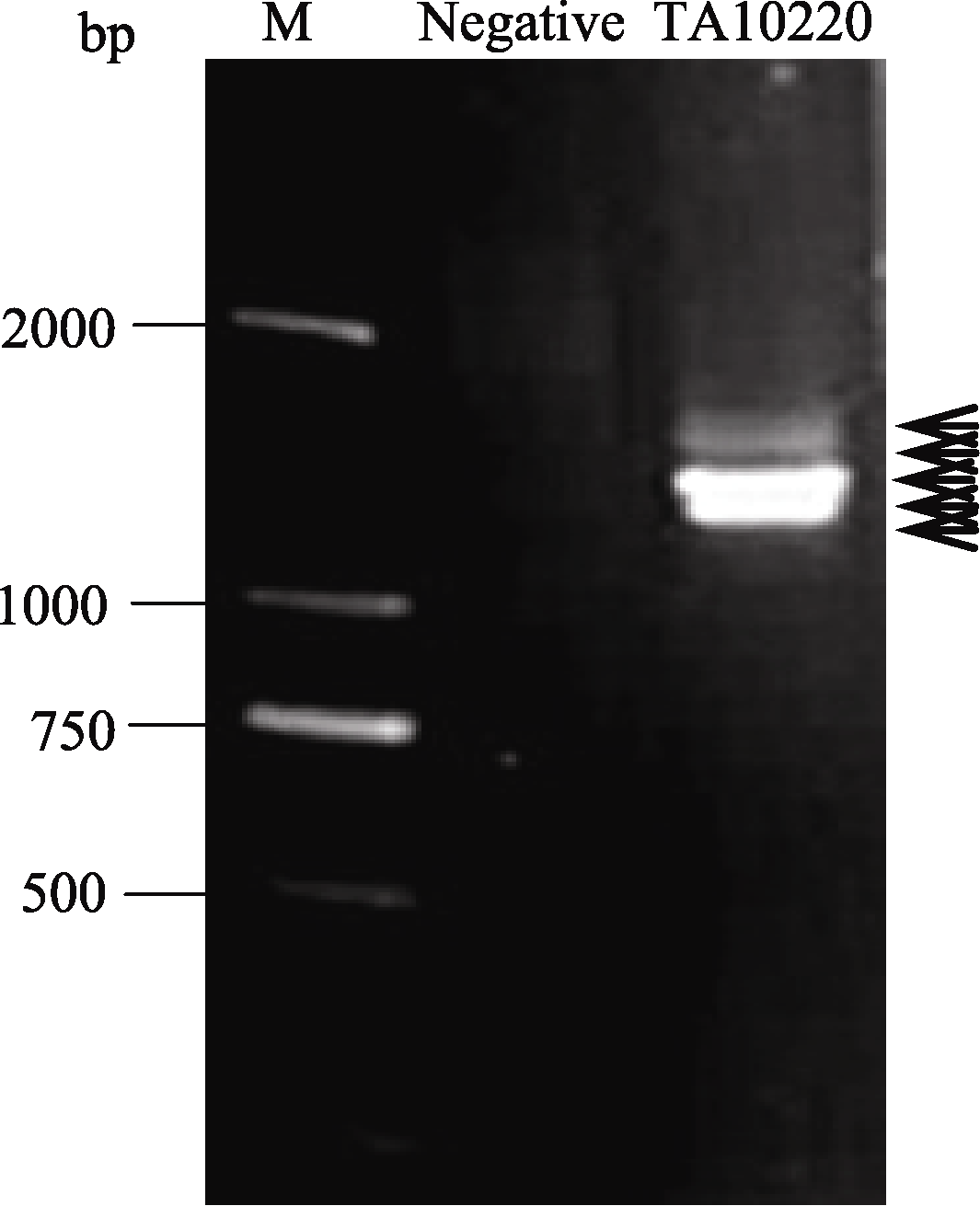 | 图1 2%琼脂糖凝胶电泳分析PCR产物M: 分子量标准DM2000; Negative: 无DNA模板的阴性对照; 箭头示目的带。Fig. 1 Amplified product separated on 2% agarose gelM: DNA marker DM2000; Negative: template DNA free; the arrows show target bands. |
用NCBI在线工具ORF Finder分析克隆获得的6条目标序列, KF887414、KF887415、KF887416和KF887417具有完整的开放阅读框(表1), 编码区内无内含子; KF887418和KF887419同样具有完整的开放阅读框, 编码区内也没有内含子的存在, 但在KF887418编码氨基酸的第323位和KF887419编码氨基酸的第269位分别出现提前终止信号(图2)。经多次测序鉴定表明, 这2个基因可能是假基因。
6条推导氨基酸序列均包括信号肽、N-端保守区、中间重复区和C-端保守区等HMW-GS典型的氨基酸序列结构特征(图2); 与小麦属A、B和D、山羊草属C和U、偃麦草属E、黑麦R和类大麦属K染色体组编码的HMW-GS氨基酸组成及结构特征相似(表2)。其中, 所有推测的氨基酸序列的信号肽均由21个氨基酸残基组成, 但在第8、第13、第16和第18氨基酸处分别出现C/L、V、L和F的多态性位点, 但SignalP分析表明, 这些多肽性位点不影
| 表1 核苷酸序列和氨基酸序列特征 Table 1 Basic characteristics of nucleotide sequence and amino acid sequence |
 | 图2 KF887414、KF887415、KF887416、KF887417、KF887418和KF887419推导氨基酸序列分析Fig. 2 Deduced amino acid sequences of KF887414, KF887415, KF887416, KF887417, KF887418, and KF887419 |
| 表2 不同来源的HMW-GS的主要结构特征的比较 Table 2 A summary of properties of the primary structure of HMW-GS from Dasypyrum villosumin comparison with some HMW-GS from common wheat and wheat-related grass |
利用MEGA5.1软件, 对来自 D. villosum的6个克隆序列与来自小麦、山羊草属、偃麦草属、黑麦、类大麦属等不同基因组和已经公布的簇毛麦属V染色体组编码的HMW-GS氨基酸序列进行全序列、N-端保守区和C-端保守区聚类分析。在全序列聚类图中, 本试验获得的6个序列与y型HMW-GS的关系较近, 与x型HMW-GS的关系较远; 在y型HMW- GS类型中, 与小麦A、B和D、山羊草属的C和U、偃麦草属E、黑麦R和类大麦属K染色体组的亲缘关系较远, 但与簇毛麦属V染色体组的HMW-GS同属一支(图3-A)。N-端保守区聚类图显示与全序列分析结果相似(图3-B), 但C-端保守区分析结果略有不同: 除KF887414外的各序列与x型HMW-GS的C-端保守区的亲缘关系较近, 其中又与簇毛麦V的关系最近, 而与小麦A、B和D、山羊草属的C和U和黑麦R染色体组的亲缘关系较远, KF887414与y型的HMW-GS的关系较近, 尤其与小麦D和山羊草属U染色体组的亲缘关系较近(图3-C)。
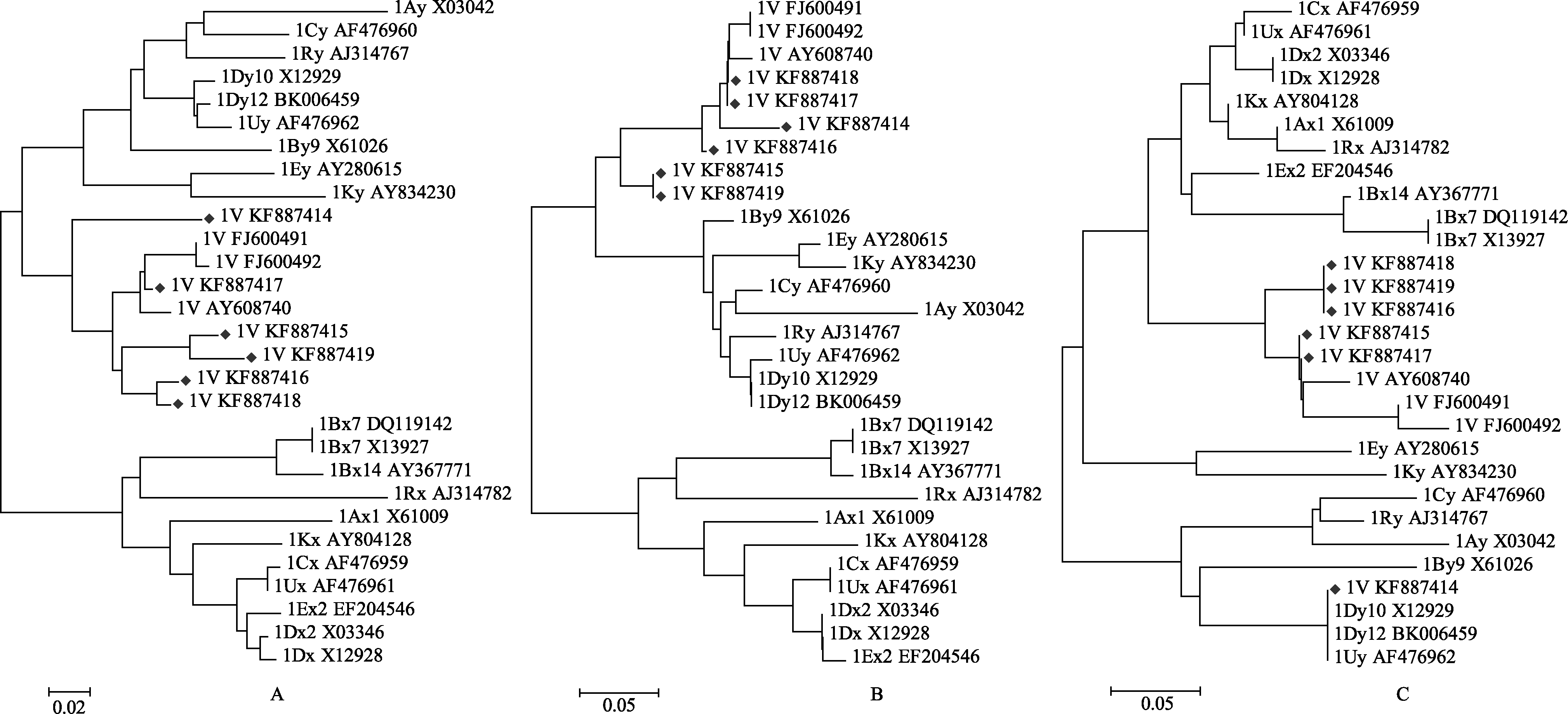 | 图3 簇毛麦HMW-GS编码的蛋白序列系统演化分析(Neighbor-joining进化树)Fig. 3 Phylogenetic analysis of the deduced amino acid sequence of HMW-GS from Dasypyrum villosum (Neighbor-joining tree) |
A: 全序列; B: N-端保守区; C: C-端保守区。
A: full length sequence; B: N-terminal region; C: C-terminal region.
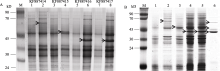
由于KF887418和KF887419是序列内部有提前终止信号的假基因, 所以未对其进行原核表达。其他4个初步判定为有功能的基因, 根据目的序列的完整编码区, 设计2对表达引物, 筛选表达基因插入方向正确的阳性克隆并测序鉴定。然后加入IPTG诱导6 h, 表达载体和能够表达的基因的重组质粒在 E. coli Rosetta-gami B(DE3) 中表达, 表达载体上的His标签的大小为2.101 kD, 融合表达蛋白产物的分子质量如表1所示, 与电泳检测的蛋白质条带大小相似(图4-A); 原核表达的蛋白质与来源于种子的高分子谷蛋白亚基具有基本相同的迁移率(图4-B), 表明克隆的HMW-GS基因在大肠杆菌中正确表达。
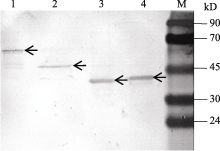
重组质粒经诱导产生的蛋白质条带与His抗体产生特异杂交信号(图5), 表明目的蛋白基因得到了成功表达。
 | 图4 KF887414、KF887415、KF887416和KF887417基因的诱导表达产物和种子HMW-GS的SDS-PAGE分析M: 蛋白质分子质量标准Marker II。A图中, 1、3、5和7泳道为未诱导重组质粒表达产物; 2、4、6和8泳道为经IPTG诱导后的重组质粒的表达产物; 箭头示基因的表达产物。B图中, 泳道1为未诱导重组质粒表达产物; 2~5泳道为经IPTG诱导后的重组质粒的表达产物, 依次是 KF887414、KF887415、KF887416和KF887417基因; 泳道6为种子提取的HMW-GS; 箭头示目的蛋白。Fig. 4 Analyses of expressed proteins of genes from KF887414 to KF887417 and comparison with seed HMW-GS using SDS-PAGEM: Protein ladder marker II. In panel A, lanes 1, 3, 5, and 7 show proteins of recombinant plasmid before induction and lanes 2, 4, 6, and 8 show proteins of recombinant plasmid after adding IPTG; the arrows show expression products. In panel B, lane 1 shows the protein of recombinant plasmid before induction; lanes 2, 3, 4, and 5 show proteins of recombinant plasmid KF887414, KF887415, KF887416, and KF887417 after adding IPTG, respectively; lane 6 shows HMW-GS extracted from Dasypyrum villosum seed; the arrows show target proteins. |
 | 图5 KF887414、KF887415、KF887416和KF887417基因的诱导表达产物Western blot检测M: 蛋白质分子质量标准Marker II。1、2、3和4泳道分别为KF887414、KF887415、KF887416和KF887417基因经IPTG诱导后的表达产物; 箭头示基因的表达产物。Fig. 5 Western blotting assay of expressed proteins of genes from KF887414 to KF887417M: Protein ladder marker II; lanes 1, 2, 3, and 4 show proteins of recombinant plasmid KF887414, KF887415, KF887416, and KF887417 after adding IPTG, respectively; the arrows show target proteins. |
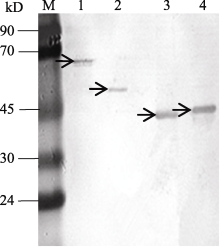
诱导后的蛋白样品经蛋白提取和处理, 用His-Trap HP柱亲和纯化目的蛋白, 回收蛋白溶液, 用12% SDS-PAGE检测, 得到单一条带, 大小与融合表达产物的相似(图6), 表明蛋白纯化成功。
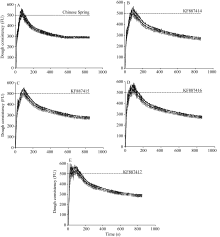
在4 g基础面粉中添加还原剂DTT和氧化剂KIO3后, 得到粉质曲线及相应参数(图7-A和表3); 利用氧化还原反应原理, 将纯化后的目的蛋白10 mg加入对照面粉, 得到粉质曲线图(图7-B~E)和相关参数(表3)。与对照相比, 簇毛麦HMW-GS的加入引
 | 图6 KF887414、KF887415、KF887416和KF887417基因的诱导表达产物柱纯化M: 蛋白质分子质量标准Marker II。1、2、3和4泳道分别为KF887414、KF887415、KF887416和KF887417基因经IPTG诱导后的表达产物; 箭头示基因的表达产物。Fig. 6 Analyses of purification of expressed proteins of genes from KF887414 to KF887417 using SDS-PAGEM: Protein ladder marker II; lanes 1, 2, 3, and 4 show proteins of recombinant plasmid KF887414, KF887415, KF887416, and KF887417 after adding IPTG, respectively; the arrows show target proteins. |
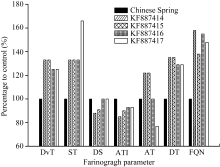
起了粉质曲线和粉质参数变化, 其中, DvT、ST、DT和FQN都有不同程度的增加, 并且增幅均存在显著或极显著差异; MTI都有不同程度的下降, 降幅差异显著或极显著; 与对照相比, 加入KF887414和KF887415基因表达产物的DS减小且差异显著, 而加入KF887416和KF887417基因表达产物的DS没有变化; KF887414和KF887415样品的AT时间推后并与对照差异显著, KF887416的AT无变化, 而KF887417的AT提前并且变化极显著(图7、图8和表3)。
 | 图7 簇毛麦来源的HMW-GS对粉质曲线的影响A: 中国春添加DTT、KIO3; B: 中国春添加DTT、KIO3和KF887414; C: 中国春添加DTT、KIO3和KF887415; D: 中国春添加DTT、KIO3和KF887416; E: 中国春添加DTT、KIO3和KF887417。Fig. 7 Effects of HMW-GS from Dasypyrum villosumon farinograph parametersA: Chinese Spring with DTT and KIO3; B: Chinese Spring with DTT, KIO3,and KF887414; C: Chinese Spring with DTT, KIO3,and KF887415; D: Chinese Spring with DTT, KIO3,and KF887416; E: Chinese Spring with DTT, KIO3,and KF887417. |
| 表3 纯化蛋白亚基的粉质参数 Table 3 Farinograph parameters of purified HMW-GS from Dasypyrum villosum |
表中数据为3次试验的平均值, 均值后大写字母表示0.01差异显著水平。
Data are the average of triplicated micro-mixing experiments. Means marked with different letters are significantly different at P < 0.01. DvT: development times; ST: stability time; DS: degree of time; MTI: mixing tolerance index; AT: arrival time; DT: departure time; FQN: farinograph quality number.
 | 图8 添加KF887414、KF887415、KF887416和KF887417基因表达蛋白的中国春与中国春(对照)粉质参数的比较DvT: 形成时间; ST: 稳定时间; DS: 弱化度; MTI: 机械耐力系数; AT: 及线时间; DT: 离线时间; FQN: 粉质质量参数。Fig. 8 Comparison of farinogragh parameters between Chinese Spring (control) and Chinese Spring with expressed proteins of KF887414, KF887415, KF887416, and KF887417DvT: development time; ST: stability time; DS: degree of time; MTI: mixing tolerance index; AT: arrival time; DT: departure time; FQN: farinograph quality number. |
利用设计的HMW-GS特异引物, 从簇毛麦TA10220基因组中获得43个阳性克隆, 鉴定出6种特异的编码序列。根据编码序列推断, KF887418和KF887419分别在氨基酸第323和第269位处出现终止密码(TGA), 推测可能是由于碱基的转换突变造成的。关于贮藏蛋白编码序列的内部出现终止密码的现象已有报道, 由密码子CAG (Gln)突变成TAG(终止密码)或由CAA (Gln)突变成TAA (终止密码)造成了1Ay、1Ax、1Dx不能正常表达, 成为了假基因, 由于碱基的转换突变引起的基因沉默同样也存在于醇溶蛋白基因中[ 30, 31, 32, 33]。Zhong和Qualset[ 34]通过提取种子全蛋白对122份来源不同的二倍体簇毛麦HMW-GS分析表明, 这些簇毛麦所编码的HMW-GS均在中国春1Dx2亚基和1Dy12亚基之间。理论上这122份簇毛麦的HMW-GS亚基分子量也在70~140 kD之间, 但是该结论未经基因水平的验证, 同时通过提取种子的全蛋白来分析HMW-GS无法排除种子醇溶蛋白和LMW-GS对结论的影响。本研究利用专门分离HMW-GS的方法提取种子HMW-GS, 通过对提取条件的优化尽可能地排除醇溶蛋白和LMW-GS的影响。虽然其余4个真基因的表达蛋白分子量均小于70 kD, 但是体外表达蛋白质和种子来源的HMW-GS具有基本相同的迁移率(图4-B), 表明本研究所得到的4条蛋白质来自簇毛麦TA10220, 并将这6个基因序列根据类型划分为1个y型亚基和5个混合型亚基。显然, 来源于二倍体簇毛麦TA10220的HMW-GS无论在亚基数目及亚基类型上都与普通小麦存在显著差异, 该物种本身的异花授粉特性所导致的高度异质可能对此产生一定的影响[ 13, 14], 而其分子机制尚需进一步研究。
HMW-GS是影响加工品质重要的遗传因素, 簇毛麦的HMW-GS多为普通小麦不具有的新亚基类型, 并已证实对面筋质量有显著的正向效应。了解簇毛麦HMW-GS基因的分子特征, 对于进一步利用新型HMW-GS基因改良普通小麦面筋质量具有一定的意义。目前已从普通小麦及其亲缘属种中克隆了众多HMW-GS基因, 这些基因的分子量在1.8~2.6 kb之间, 编码的蛋白质分子量在60~140 kD之间(表2)。随着研究的深入以及对结构功能的认识, 发现一些小于1.8 kb的麦谷蛋白基因同样具有HMW-GS的特点[ 21, 22, 23, 24, 35, 36, 37, 38, 39, 40, 41, 42]。本文利用同源克隆技术克隆到的簇毛麦HMW-GS基因小于1.8 kb, 但其推导氨基酸序列, 具有HMW-GS结构特点(图2), 编码区没有内含子, 具有21个氨基酸残基的信号肽, 高度保守的N-和C-端及中间重复区[ 11, 12], 尽管分子量较小, 也应该是HMW-GS基因。这与前人所克隆的簇毛麦HMW-GS基因结果[ 20, 21, 22, 23, 24]较为一致。由于簇毛麦与小麦的亲缘关系较远, 传统的以大小区分HMW-GS的方法, 在簇毛麦研究分析上也许存在一定的局限性。普通小麦HMW-GS大小的分类方法不适合对远缘材料的分类(表2), 即不能仅仅依据SDS- PAGE上的迁移率区分。尤其在对利用簇毛麦等远缘材料所创制的材料, 包括易位系、附加系鉴定的时候, 不能仅仅靠蛋白质分子量大小进行染色体易位片段的判断。
本研究得到的所有簇毛麦HMW-GS基因编码的氨基酸序列, N-端保守区有5个半胱氨酸残基, 并且第3和第4位半胱氨酸残基相邻, 与报道的y型HMW-GS的N-端保守区的特征一致(图2和表2); N-进化树分析也表明该6条推导的氨基酸序列的N-端保守区与y型关系较近与x型关系较远(图3-B)。KF887414的中间重复区六肽(PGQGQQ)和九肽(GYYPTSP/LQQ)高度重复, 不存在三肽(GQQ), 表明KF887414基因编码的氨基酸序列具有y型HMW-GS的中间重复区的特征[ 2, 3], 而KF887415~ KF887419除重复出现六肽(PGQGQQ)和九肽(GYYPTSP/LQQ)外, 还具有类似三肽(GQQ)PGQ, 表明这些序列的中间重复区具有典型的x型HMW-GS特征[ 2, 3], 庞玉辉等[ 21]报道的N-端保守区具有y型HMW-GS特征的簇毛麦高分子谷蛋白的中间重复区也存在PGQ、GQQ和RGQ等三肽。本研究中, 氨基酸序列的C-端保守区具有一个半胱氨酸残基。C-端进化树分析表明KF887414与y型亲缘关系较近, 而KF887415~KF887419与x型关系较近(图3-C)。KF887414的近C-端重复区多1个半胱氨酸残基, KF887419的近N-端重复区多出1个(图2)。以上分析表明, 某些簇毛麦HMW-GS兼有普通小麦来源的x和y型的一些结构特征, 此种现象可能是簇毛麦物种本身的特异性所致。因此, 用以区分栽培小麦x和y型HMW-GS的分类方法需要进行适当完善方能反映其亲缘关系较远的野生种属所具有的同源亚基的属性。
利用微量掺粉试验可以直观地检测亚基的品质效应, 簇毛麦TA10220来源的4个HMW-GS的掺入均使整个面团的空间结构经历了某些剧烈改变, 并最终表现为粉质曲线图的明显变化(图7): 面团的形成时间延长, 表明上述亚基具有提高基础面粉面筋质量的潜力; 稳定时间延长表明提高了面团的耐揉性、韧性和面筋的强度; 离线时间延长表明面筋强度或耐受过度搅拌的能力增强; 粉质质量参数显著提高表明面团的综合质量具有显著的正向效应; 耐搅拌指数降低说明面团的耐揉性提高(图8和表3)。因此, 簇毛麦中的HMW-GS对面团的品质有正向效应。通过杂交育种的方式将簇毛麦的HMW-GS基因转入小麦, 对改良普通小麦的加工品质, 增加其遗传多样性具有重要意义。
克隆到6个HMW-GS基因序列, 均小于1.8 kb, 其中KF887418和KF887419是编码区具有终止信号的假基因, 这6个序列均具有HMW-GS显著的结构特征, 并且与已知的HMW-GS的亲缘关系较近; 其中KF887414为y型HMW-GS, 而其他序列均同时具有x和y型HMW-GS结构特征。簇毛麦来源的HMW-GS对面粉的加工品质有显著的正向效应。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|