谷子m6A甲基转移酶基因SiMTA1的启动子序列特征和基因表达模式分析
沈傲, 刘敏, 倪迪安, 刘炜
作物学报
2025, 51 ( 7):
1969-1978.
DOI: 10.3724/SP.J.1006.2025.44210
MTA作为植物m6A甲基转移酶之一, 主要参与RNA的甲基化修饰, 同时还可与其他酶相互作用, 影响胚胎发育, 在植物的生长发育中扮演着关键角色。本研究通过拟南芥甲基转移酶序列同源比对, 得到谷子SiMTA1基因(accession no. PQ801843), 该基因DNA序列全长4239 bp, CDS序列2121 bp, 包含706个氨基酸残基。对其核苷酸及蛋白序列进行生物信息学分析、并解析其启动子顺式作用元件, 通过qRT-PCR方法对SiMTA1在时空表达和各种胁迫及激素处理下的表达模式进行研究。结果显示, SiMTA1具有MT-A70结构域, 即N6-腺苷-甲基转移酶(MTase)的S-腺苷甲硫氨酸(SAM)结合亚基; 二级结构预测显示, SiMTA1主要是由无规则卷曲和α-螺旋组成; SiMTA1启动子包含多种胁迫和植物激素信号应答元件; SiMTA1在孕穗期的茎节部的表达量较高; 盐、干旱、生长素、细胞分裂素等均可诱导SiMTA1表达。本研究通过对谷子中甲基转移酶基因SiMTA1启动子特性及其在正常及胁迫条件下时空表达模式的研究发现, SiMTA1可能在谷子发育及胁迫和植物激素响应过程中发挥作用。研究结果为后续谷子耐逆品种的改良及新品种培育提供了理论依据及候选基因资源。
| 引物名称Primer name | 引物序列Primer sequence (5'-3') | | Q-SiMTA1-F1 | TGGCTGACGATGAAATGAGG | | Q-SiMTA1-R1 | GCCAGTTCGGATGATACGTT |
View table in article
表1
本研究所用引物
正文中引用本图/表的段落
RNA修饰是一种在转录后阶段发生的核酸修饰, 近年来已成为表观遗传学研究的热点[1]。N6-甲基腺嘌呤(N6-methyladenine, m6A)是一种常见的核酸修饰, 特别是在真核生物的mRNA中, 这种修饰涉及将一个甲基团(-CH3)添加到腺嘌呤碱基的N6位点上。m6A是真核生物mRNA上最丰富的化学修饰, 并且在调控基因表达、mRNA的剪接、出核、稳定性以及翻译效率等多个方面发挥着重要作用[2]。早在1979年, Kennedy和Lane[3]首次在小麦中发现N6-甲基腺嘌呤(m6A)的存在, 开启了植物中关于m6A修饰相关酶的研究。随后, Haugland和Cline [4]在1980年发现燕麦胚芽鞘的mRNA中也含有m6A。2014年, Luo等[5]首次对拟南芥中与mRNA甲基化相关的基因进行系统性鉴定, 并提出m6A修饰的沉积与mRNA丰度呈正相关。Dominissini等[6-7]发现, m6A修饰广泛分布于mRNA的各个区域, 包括5'UTR、CDS、3'UTR、终止密码子以及转录起始位点附近, 且在终止密码子和3'UTR区域的富集程度较高。在植物中, m6A甲基化过程是由特定的甲基转移酶介导, 在拟南芥中已经鉴定出几种关键的m6A甲基转移酶, 包括MTA (Methyltransferase A)、MTB (Methyltransferase B)、FIP37 (FKBP12 interacting protein 37 kD)和VIR (Virilizers)等[8]。其中, MTA是拟南芥中最早被发现的甲基转移酶之一, 具有m6A催化活性, 并与FIP37共同构成植物m6A甲基转移酶复合物的核心组成部分, 在调控芽、根生长以及叶和花的发育中扮演关键角色[9-10]。研究表明, 在拟南芥中, m6A甲基转移酶基因MTA和FIP37受冷热胁迫时表达量略有上升; 而在寒冷条件下水稻中OsMTA、OsMTB和OsVIR的表达量则降低[11]。在干旱条件下, m6A甲基化酶基因的表达对大多数植物的影响并不显著, 但在水稻中OsMTA、OsMTB和OsVIR的表达量因干旱而降低, 而杨树PtrMTA的过表达则显著提高了杨树毛状体的密度, 促进了根系的发展, 增强了杨树对干旱的适应能力[11-12]。m6A甲基化在拟南芥的盐胁迫响应中起着关键作用, MTA、MTB和VIR等突变体均表现出对盐分的敏感性[13]。此外, 也有研究表明, 植物m6A甲基化相关蛋白对外源植物生长调节剂有响应。在拟南芥中, 当ALKBH6被敲除时, 它的种子在受到ABA刺激的时候会比正常情况下更早地开始萌发, 然而它们的幼苗和根部发育并没有显示出明显的不同于野生型的特征[14]。
根据谷子甲基转移酶基因SiMTA1的CDS序列, 通过NCBI数据库设计特异性引物(表1)。由青岛擎科生物公司合成引物。以反转录获得的单链cDNA为模板, 以谷子持家基因SiActin (NCBI登录号: XM-004978702)为内参, 利用表1中特异性引物, 根据诺唯赞ChamQ Universal SYBR qPCR Master Mix试剂说明书在荧光定量PCR仪上进行目的片段的扩增。扩增体系为20 μL, 含Q-SiMTA1- F1 0.4 μL、Q-SiMTA1-R1 0.4 μL、稀释5倍的cDNA模板1 μL、2×ChamQ Universal SYBR qPCR Master Mix 10 μL、ddH2O 8.2 μL。扩增条件: 95℃ 30 s; 95℃ 10 s, 60℃ 30 s, 40个循环; 95℃ 15 s, 60℃ 60 s, 95℃ 15 s。采用2-ΔΔCt法计算SiMTA1基因在各发育阶段及不同胁迫处理下的相对表达量; 利用Excel、Origin等软件分析数据并作图。
利用拟南芥已知的MTA序列, 检索谷子基因组序列, 获得谷子中的MTA候选基因。将谷子SiMTA1与拟南芥(AtMTA)、水稻(OsMTA)、小麦(TaMTA)、玉米(ZmMTA)的MTA类蛋白进行同源比对发现, SiMTA1与其他物种的MTA蛋白在第460~650个氨基酸之间的区域显示出较高的保守性。表明在这个区域内, 各个物种的MTA蛋白可能具有相似的结构和功能。在所有比较的物种中, 玉米(ZmMTA)与谷子SiMTA1的同源性最高, 达到92.33%, 推测两者的亲缘关系较近, 可能具有相似的生物学功能(图1)。
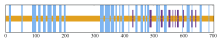
启动子SiMTA1-P正义链包含的顺式作用元件预测结果如图5, 共包含15种元件且大多位于正义链的前半段。对启动子SiMTA1-P正反义链顺式作用元件分析显示(表2), 其包含多种经典的胁迫响应和激素调控元件。其中, ABRE、DRE和LTR是典型的胁迫响应元件。含有ABRE基序的基因通常在ABA的调控下表达, 从而参与植物的逆境适应过程; DRE与植物的干旱、低温和高盐胁迫响应有关; LTR与基因的低温响应有关。此外, 启动子中还存在其他重要的功能元件, 例如AS-1参与调节生长素应答基因的表达, 影响植物的发育过程; MYC和CCGTCC-box等元件在植物应对逆境和激素信号传递中起着关键作用。顺式作用元件的多样性与其所调控的下游基因具有逆境适应及激素信号传递的生物学功能密切相关。
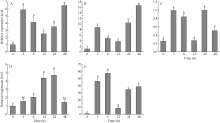
在谷子的不同发育期, 以SiMTA1在三叶期的相对表达量为参照, SiMTA1在孕穗期的表达量最高, 约为三叶期的60倍, 其次在拔节期和开花期的相对表达量也较高, 约为三叶期的18倍(图6-A)。对孕穗期SiMTA1的组织表达模式分析发现, 以基因在旗叶的相对表达量为参照, SiMTA1在茎节的表达量最高, 约为旗叶的78倍, 其次在幼穗和茎中有较高的表达, 分别为旗叶的54倍和42倍(图6-B)。
A: SiMTA1在谷子各发育时期下的相对表达量; B: SiMTA1在孕穗期不同组织的相对表达量。不同小写字母表示在0.05水平差异显著。
A: SiMTA1在盐胁迫下的相对表达量; B: SiMTA1在干旱胁迫下的相对表达量; C: SiMTA1在4℃胁迫下的相对表达量。不同小写字母表示在0.05水平差异显著。
不同激素处理下, SiMTA1基因也表现出不同的响应模式。在生长素(IAA)、细胞分裂素(6-BA)和生长素类激素(NAA)的处理下, SiMTA1基因表达呈先升高再降低再升高的变化趋势, 且基因表达升高幅度相对较高(图8-A, B, E); 在赤霉素(GA3)及乙烯(ACC)的处理下, SiMTA1的表达也呈先增加后下降的动态趋势, 但整体表达升降幅度不大(图8-C, D)。综上, SiMTA1基因对生长素类激素及细胞分裂素的响应较为明显, 随处理时间的延长具有较明显的表达模式变化, 推测SiMTA1基因可能在植物生长发育调节中发挥作用。
A: SiMTA1在生长素处理下的相对表达量; B: SiMTA1在细胞分裂素处理下的相对表达量; C: SiMTA1在赤霉素处理下的相对表达量; D: SiMTA1在乙烯处理下的相对表达量; E: SiMTA1在生长素类激素处理下的相对表达量。不同小写字母表示在0.05水平差异显著。
在基因功能研究中, 启动子区域的关键调控元件分析是重要策略之一[28]。通过研究启动子对报告基因表达的调控作用, 可以揭示其对下游基因表达和生物学功能的影响。例如, 杨春霞等[29]通过含有rd29A启动子驱动的GUS基因表达载体导入烟草, 发现在逆境处理下GUS基因的表达量显著提升, 这表明rd29A启动子可以被逆境条件激活。姜骁等[30]构建以NTORK1启动子为控制序列的GUS基因瞬时表达载体, 发现该启动子能够响应高剂量的SA和低剂量的NAA。本研究基于Plant CARE数据库对谷子SiMTA1基因上游启动子区域进行分析, 发现该区域含有大量参与胁迫响应和激素响应的顺式作用元件, 其中响应逆境胁迫的元件有8个, 响应植物激素的元件有5个, 共同响应的元件有3个, 说明SiMTA1-P启动子可能具有多重调控活性, 进一步推测, 其下游基因SiMTA1可能在谷子的胁迫适应及激素信号传导中发挥重要作用。
基因表达模式的分析是研究基因在不同条件下(如组织类型、发育阶段或环境胁迫下)表达特性的重要手段。为深入探究SiMTA1在时空表达和各种胁迫下的表达特性,本研究采用qRT-PCR技术分析SiMTA1基因在谷子不同发育时期及各组织中的表达情况, 及其在高盐、干旱、低温、生长素、细胞分裂素、乙烯、赤霉素以及生长素类激素处理下的表达模式。结果显示, SiMTA1基因在孕穗期的表达量较高, 特别在该时期的茎节部位, 这种表达模式与拟南芥、玉米中的MTA基因的功能类似。在拟南芥中, 敲除MTA基因会导致种子停留在球形胚阶段无法进一步发育[31], 通过特定启动子(如ABI3)驱动MTA的表达, 可以恢复部分MTA突变体种子的早期胚胎发育; 但在后期停止表达MTA后, 植株的m6A水平会显著下降[32]。在玉米中, ZmMTA功能丧失会导致玉米胚胎和胚乳发育停滞, 且m6A修饰基因的5'区CHH甲基化显著降低[33]。以上结果表明, MTA基因参与植物的生长过程并在谷子的生长发育过程中起重要作用。同时SiMTA1在各种胁迫处理下均有表达, 其中在盐处理下其表达呈上升趋势, 盐处理48 h时达到峰值, 这表明SiMTA1与拟南芥中的MTA基因类似, 均对盐胁迫表现出显著的响应。此外PEG模拟干旱和4℃低温处理下, SiMTA1基因的表达量呈上升和波动的趋势, 相比之下, 水稻中的OsMTA基因在寒冷和干旱条件下表达量均降低, 表明MTA基因在逆境胁迫下可能通过表达来响应环境变化。谷子幼苗经生长素、细胞分裂素和生长素类激素处理后, SiMTA1基因的表达量均表现出先升高随后下降然后再次升高的变化趋势, 而乙烯处理下SiMTA1基因的表达量表现为先升高后降低, 赤霉素处理后SiMTA1基因的表达量呈波动变化。以上结果显示, SiMTA1基因对多种胁迫和激素处理下可产生响应, 其响应情况与该基因启动子SiMTA1-P中所包含的多种胁迫和激素响应元件所具有的对下游基因功能的调控功能相一致。表明谷子在受到干旱、盐等胁迫和激素处理下, 启动子可对SiMTA1基因的转录和表达发生调控, 从而影响基因的表达量, 使植物对逆境胁迫做出响应以减少胁迫对植物的伤害, 基因也进一步参与到植物激素响应和激素信号传导等过程。
本文的其它图/表
|