小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1 ] 。受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2 ] 。在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2 ] 。然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病。发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一。
研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] 。其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] 。利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高。黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定。稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程。本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持。
1 材料与方法
1.1 回交群体的构建
周麦16是黄淮冬麦区南片的主栽品种之一, 也是近年来该麦区应用最广泛的骨干亲本。利用周麦16与矮败小麦的不育株杂交, 并用周麦16作为轮回亲本与后代中的不育株连续回交, 培育出矮败周麦16。矮败周麦16后代总是有一半数量的高秆可育株和一半数量的矮秆雄性不育株。可育株与周麦16相似, 不育株除株高较矮和雄性不育以外, 其他性状与轮回亲本周麦16相似。2014年, 利用矮败周麦16的不育株作母本, 与携带Fhb1 基因的中抗赤霉病小麦品种(系)宁麦9号、生选6号、建阳798、建阳84、苏麦3号和宁麦13杂交, 产生6个F1 组合。2015年, 利用周麦16作父本, 分别与6个F1 组合中的不育株回交, 获得BC1 F1 种子。2016年, 采用Fhb1 基因的KASP标记检测6个BC1 F1 群体, 对携带Fhb1 基因的不育株继续回交, 选择携带Fhb1 基因的可育株用于赤霉病抗性鉴定。
淮麦20是黄淮冬麦区抗赤霉病鉴定的中感对照, 在本研究中作为抗赤霉病人工接种鉴定和田间病圃自然鉴定的中感赤霉病对照。另外, 在人工接种鉴定中, 扬麦158和安农8455分别作为中抗赤霉病对照和高感赤霉病对照。
1.2 赤霉病抗性表型评价
2016年10月, 将来自上述6个回交群体携带Fhb1 基因的11个可育株和11个不携带Fhb1 基因的可育株、轮回亲本周麦16、中感赤霉病对照品种淮麦20、中抗赤霉病对照品种扬麦158和高感赤霉病的对照品种安农8455种植在江苏省农业科学院(南京市)赤霉病鉴定圃, 采用单花滴注法[17 ] 人工接种鉴定。采用完全随机区组设计, 单行区, 行长2.0 m, 行距0.3 m, 株距5.0 cm, 2次重复。单花滴注的赤霉菌孢子液10 μL, 孢子浓度为5 × 105 mL-1 。在始花期, 每个株系接种10穗, 套袋保湿72 h后, 在弥雾保湿条件下继续生长, 接种后21 d调查病小穗数, 作为赤霉病抗性评价指标[17 ] 。
2016年11月, 将上述用于人工接种鉴定的所有可育株、轮回亲本、中感对照品种淮麦20种植在福建省南平市农业科学研究所赤霉病田间鉴定圃进行自然鉴定。采用完全随机区组设计, 单行区, 行长1.0 m, 行距0.3 m, 株距5.0 cm, 3次重复。扬花后20 d, 每个材料每重复调查15穗。采用病情指数作为田间病圃鉴定的评价指标[18 ] 。
利用Microsoft Excel 2007数据分析工具对携带和不携带供体Fhb1 基因的回交后代病小穗数和病情指数进行t 测验。利用SAS v9.2软件(SAS Institute)对不同Fhb1 供体的回交后代间病小穗数和病情指数分别进行多重比较分析(LSD法)。
1.3 Fhb1 基因的KASP标记检测
利用DNA快速提取试剂盒(北京天根生化科技有限公司)提取小麦叶片的基因组DNA。采用BIO-RAD S1000 Thermal Cycler PCR System (Bio-Rad Biotechnology Laboratory, 美国) PCR扩增仪, 5 μL PCR体系包括2.5 μL KASP Master Mix (Laboratory of the Government Chemist, 英国), 正反引物0.056 μL (100 μmol L-1 ), Mg2+ 0.04 μL (50 mmol L-1 ), DNA模板(20~50 ng μL-1 ) 2.2 μL, 0.205 μL去离子H2 O。
采用降落PCR扩增, 程序为95°C热启动15 min, 9个降落循环(95°C 20 s, 65°C 20 s, 从65°C每个循环降低1°C), 95°C变性10 s, 57°C退火60 s, 35个循环, 延伸5 min [19 ] 。反应结束后, 在酶标仪SYNERGY H1MF microplate reader (BioTek Instruments, Inc., 美国)上读取荧光数据, 并导入Kluster Caller software version 2.24.0.11 (http://kbioscience. co.uk/)作基因分型。
2 结果与分析
2.1 回交后代基因型分析
利用Fhb1 基因KASP标记对供体亲本宁麦9号、生选6号、建阳798、建阳84、苏麦3号、宁麦13和轮回亲本周麦16进行基因型检测, 6个供体亲本全部含有Fhb1 基因, 而轮回亲本周麦16不含有Fhb1 基因。同时, 利用该标记对6个BC1 F1 回交群体共650个不育株和349个可育株进行基因型分析, 不育株共检测出60株携带Fhb1 基因, 这些不育株用作母本继续与轮回亲本回交; 可育株共检测出40株含有Fhb1 基因, 从中随机选取种子量充足的11个可育株用于赤霉病抗性人工接种和自然病圃鉴定。
2.2 同一供体回交后代赤霉病抗性比较
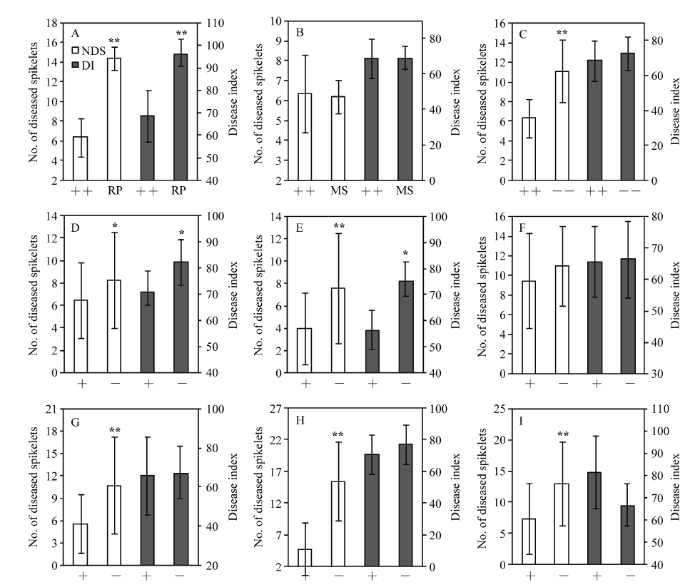
为了比较同一供体Fhb1 基因对赤霉病抗性的影响, 以及在同一遗传背景下来自不同供体的Fhb1 基因对赤霉病抗性的影响, 分别从上述相应的6个回交群体中选取11株携带Fhb1 基因的可育株和11株不携带Fhb1 基因的可育株, 按株系种植, 进行赤霉病抗性对比分析, 同时对携带Fhb1 基因的可育株与轮回亲本周麦16、中感赤霉病对照淮麦20、高感赤霉病对照安农8455及中抗对照扬麦158的赤霉病抗性进行比较。人工接种鉴定的发病情况比较充分(图1 ), 来自6个供体携带Fhb1 基因的11个株系的平均病小穗数和平均病情指数分别比轮回亲本周麦16小8.1 (P < 0.01)和28.4 (P < 0.01); 与不携带Fhb1 的11个株系相比, 平均病小穗数和平均病情指数分别少4.8 (P < 0.01) 和4.0 (图2 )。整体而言, 11个携带Fhb1 基因株系的赤霉病抗性达到中感对照淮麦20水平。单花滴注接种鉴定结果表明, 除建阳798外的5个供体回交群体中携带Fhb1 基因的株系, 病小穗数均显著低于不含有Fhb1 基因的株系(P < 0.05或P < 0.01)。5个供体回交群体的田间病圃鉴定结果与人工接种鉴定结果趋势一致, 仅宁麦13表现不一致。其中, 携带宁麦9号和生选6号Fhb1 基因的回交后代株系病情指数显著低于不携带Fhb1 的回交后代株系(P < 0.05)。
图1 单花滴注接种鉴定携带和不携带供体Fhb1 基因的回交后代、轮回亲本及对照赤霉病抗性表现Fhb1 基因回交后代的病小穗数明显少于不携带供体Fhb1 基因的回交后代。轮回亲本周麦16较中感对照淮麦20 (G)、高感对照安农8455较中抗对照扬麦158 (H)具有更多的病小穗数。+: 携带Fhb1 基因; –: 不携带Fhb1 基因; RP: 轮回亲本; MS: 中感; HS: 高感; MR: 中抗。
Fig. 1 Performance of FHB resistance in the backcrossing progenies with or without Fhb1 gene from different donors, recurrent parent and controls using the floret-inoculation methodFhb1 gene from donors Ningmai 9 (NM 9, A), Shengxuan 6 (SX 6, B), Jianyang 798 (JY 798, C), Jianyang 84 (JY 84, D), Sumai 3 (SM 3, E), and Ningmai 13 (NM 13, F) were obviously lower than the backcrossing progenies without Fhb1 gene. The recurrent parent Zhoumai 16 (ZM 16) and the highly susceptible control Annong 8455 (AN 8455) had more number of diseased spikelets than Huaimai 20 (HM 20), the moderately susceptible control (G) and Yangmai 158 (YM 158), the moderately resistant control (H). +: Fhb1 genotype; –: non-Fhb1 genotype; RP: recurrent parent; MS: moderately susceptible; HS: highly susceptible; MR: moderately resistant.
图2 携带不同供体Fhb1 基因回交后代与轮回亲本、中感对照及不携带Fhb1 基因回交后代间病小穗数和病情指数比较Fhb1 基因的所有回交后代与轮回亲本周麦16 (A)、中感对照淮麦20 (B)和不携带Fhb1 基因的所有回交后代(C)病小穗数和病情指数比较。携带宁麦9号Fhb1 基因的回交后代(D)、携带生选6号Fhb1 基因的回交后代(E)、携带建阳798 Fhb1 基因的回交后代(F)、携带建阳84 Fhb1 基因的回交后代(G)、携带苏麦3号Fhb1 基因的回交后代(H)和携带宁麦13 Fhb1 基因的回交后代(I)分别与其不携带Fhb1 基因的回交后代间病小穗数和病情指数比较。NDS: 病小穗数(人工接种); DI: 病情指数(自然发病)。++: 携带Fhb1 基因的所有回交后代; RP: 轮回亲本; MS: 中感对照; ––: 不携带Fhb1 基因的所有回交后代; +: 携带Fhb1 基因的回交后代; –: 不携带Fhb1 基因的回交后代。误差线上*和**分别表示在P < 0.05和P < 0.01水平下差异显著。
Fig. 2 Comparison of number of diseased spikelets and disease index between backcrossing progenies with Fhb1 gene from different donors and recurrent parent, moderately susceptible control and the backcrossing progenies without Fhb1 gene Fhb1 backcrossing progenies and the recurrent parent Zhoumai 16 (A), the moderately susceptible control Huaimai 20 (B) and all non-Fhb1 backcrossing progenies (C), and between non-Fhb1 and Fhb1 backcrossing progenies from the donor cultivars Ningmai 9 (D), Shengxuan 6 (E), Jianyang 798 (F), Jianyang 84 (G), Sumai 3 (H) and Ningmai 13 (I). NDS: number of diseased spikelets (floret-inoculation method); DI: disease index (natural infection nursery); ++: all Fhb1 backcrossing progenies; RP: recurrent parent; MS: moderately susceptible control; ––: all Fhb1 backcrossing progenies; +: Fhb1 backcrossing progenies; –: non-Fhb1 backcrossing progenies. * and ** above the error bars indicate significant difference at P < 0.05 and P < 0.01, respectively.
2.3 不同供体回交后代赤霉病抗性比较
尽管携带来自于6个供体Fhb1 基因的回交后代对赤霉病均表现出一定的抗性, 但是不同供体回交后代在抗病性上表现出显著差异。根据单花滴注接种法鉴定结果, 抗病性由强到弱依次为生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798的回交后代(表1 ), 生选6号回交后代对赤霉病抗性接近中抗水平; 根据田间病圃自然发病鉴定结果, 抗病性由强到弱排序为生选6号、建阳798、建阳84、苏麦3号、宁麦9号和宁麦13回交后代。两种鉴定方法的鉴定结果均表明, 生选6号的回交后代抗病性表现最好。
3 讨论
矮败小麦具有雄性败育彻底、不育性稳定、异交结实率高的优点, 同时, 由于显性矮秆基因Rht-D1c 与显性雄性不育基因Ms2 紧密连锁, 矮秆不育株与正常株高的可育株很容易区别, 简化了可育株和不育株的人工鉴定和标记过程, 是理想的小麦遗传改良工具[20 ,21 ] 。我们把矮败小麦的Rht-D1c 和Ms2 基因回交转育到黄淮冬麦区推广品种和骨干亲本, 培育出了这些品种和骨干亲本的近等基因系, 例如, 矮败周麦16、矮败郑麦9023、矮败济麦22、矮败轮选987等。本研究利用矮败周麦16与具有Fhb1 基因的品种(系)杂交和回交, 利用KASP标记对后代进行Fhb1 基因型分析, 对携带Fhb1 基因的矮秆不育株, 继续与黄淮麦区的品种(系)回交; 对携带Fhb1 基因的可育株进行赤霉病抗性鉴定和农艺性状选择, 培育符合黄淮冬麦区生态类型、赤霉病抗性得到显著改良的新品种。这种策略发挥了矮败小麦杂交方便、回交后代群体大的优势, 有利于较快地选择出既具有轮回亲本优良的农艺性状、赤霉病抗性又得到改良的新品种。
抗赤霉病基因Fhb1 位于3B染色体Xgwm533.1 - Xgwm493 标记区间内, 抗性等位变异来自苏麦3号[4 ,5 ] 。陆维忠[22 ] 采用4个赤霉病抗病品种与2个感病品种构建的6个群体进行抗赤霉病QTL发掘证实, 抗病品种携带的主效QTL均在3B染色体Xgwm533.1 - Xgwm493 之间, 单个标记的选择效率达到70%。在此基础上, Liu等[23 ] 进一步将Fhb1 定位在231 kb区间, 并开发了共显性特异标记UMN10, 其选择效率明显高于SSR标记。在本研究中, 利用Fhb1 的功能KASP标记鉴定发现, 含有Fhb1 基因的品种一般对赤霉病的抗性较好, 例如, 中抗赤霉病品种(系)生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798均含有Fhb1 基因; 同时也发现有些赤霉病抗性较好的品种不含有Fhb1 基因(例如扬麦158), 但未发现既具有该标记又高感赤霉病的品种。这一方面说明Fhb1 基因对赤霉病抗性的有效性, 另一方面也说明除Fhb1 基因外, 有些品种可能携带其他抗赤霉病基因。
在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因。目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] 。与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平。与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平。
尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异。田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强。生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当。分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] 。本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究。宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] 。与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关。
4 结论
6个回交群体中携带Fhb1 基因的回交后代在平均病小穗数和平均病情指数上均显著低于轮回亲本周麦16和不含有Fhb1 基因的回交后代, 赤霉病抗性达到中感水平。不同供体回交后代在抗病性上表现出明显差异, 其中最好的是生选6号。通过Fhb1 基因分子标记辅助选择技术改良黄淮冬麦区小麦品种的赤霉病抗性是可行的。
致谢 感谢中国农业科学院作物科学研究所夏先春博士在KASP基因分型方面提供的技术支持。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献
文献选项
[1]
Yin Y Liu X Li B Ma Z Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F
. graminearum collected from wheat in China. Phytopathology , 2009 , 99 : 487 -497
[本文引用: 1]
[2]
程顺和 , 张勇 , 别同德 , 高德荣 , 张伯桥 . 中国小麦赤霉病的危害及抗性遗传改良
. 江苏农业学报 , 2012 , 28 : 938 -942
[本文引用: 2]
Cheng S H Zhang Y Bie T D Gao D R Zhang B Q Damage of wheat Fusarium head blight epidemics and genetic improvement of wheat for scab resistance in China
.Jiangsu J Agric Sci 2012 , 28 : 938 -942 (in Chinese with English abstract)
[本文引用: 2]
[3]
刘新月 , 裴磊 , 卫云宗 , 张正斌 , 高辉明 , 徐萍 . 气温变化背景下中国黄淮旱地冬小麦农艺性状的变化特征——以山西临汾为例
. 中国农业科学 , 2015 , 48 : 1942 -1954
[本文引用: 1]
Liu X Y Pei L Wei Y Z Zhang Z B Gao H M Xu P Agronomic traits variation analysis of Huanghuai dryland winter wheat under temperature change background in China: taking Linfen, Shanxi as an example
.Sci Agric Sin 2015 , 48 : 1942 -1954 (in Chinese with English abstract)
[本文引用: 1]
[4]
Anderson J A Stack R W Liu S Waldron B L Fjeld A D Coyne C Moreno-Sevilla B Fetch J M Song Q J Cregan P B Frohberg R C DNA markers for Fusarium head blight resistance QTLs in two wheat populations
.Theor Appl Genet 2001 , 102 : 1164 -1168
[本文引用: 3]
[5]
Cuthbert P A Somers D J Thomas J Cloutier S Brulé-Babel A Fine mapping Fhb1 , a major gene controlling Fusarium head blight resistance in bread wheat(Triticum aestivum L.)
. Theor Appl Genet , 2006 , 112 : 1465 -1472
[本文引用: 2]
[6]
Cuthbert P A Somers D J Brulé-Babel A Mapping of Fhb2 on chromosome 6BS: a gene controlling Fusarium head blight field resistance in bread wheat(Triticum aestivum L.)
. Theor Appl Genet , 2007 , 114 : 429 -437
[本文引用: 1]
[7]
Qi L L Pumphre M O Friebe B Chen P D Gill B S Molecular cytogenetic characterization of alien introgressions with gene Fhb3 for resistance to Fusarium head blight disease of wheat
. Theor Appl Genet , 2008 , 117 : 1155 -1166
[本文引用: 1]
[8]
Guo J Zhang X Hou Y Cai J Shen X Zhou T Xu H Ohm H W Wang H Li A Han F Wang H Kong L High-density mapping of the major FHB resistance geneFhb7 derived from Thinopyrum ponticum and its pyramiding with Fhb1 by marker-assisted selection
. Theor Appl Genet , 2015 , 128 : 2301 -2316
[本文引用: 1]
[9]
Xue S Li G Jia H Xu F Lin F Tang M Wang Y An X Xu H Zhang L Kong Z Ma Z Fine mapping Fhb4 , a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.)
. Theor Appl Genet , 2010 , 121 : 147 -156
[本文引用: 1]
[10]
Xue S Xu F Tang M Zhou Y Li G An X Lin F Xu H Jia H Zhang L Kong Z Ma Z Precise mapping Fhb5 , a major QTL conditioning resistance to Fusarium infection in bread wheat ( Triticum aestivum L.)
. Theor Appl Genet , 2011 , 123 : 1055 -1063
[本文引用: 1]
[11]
刘易科 , 佟汉文 , 朱展望 , 陈泠 , 邹娟 , 张宇庆 , 焦春海 , 高春保 . 小麦赤霉病抗性改良研究进展
. 麦类作物学报 , 2016 , 36 : 51 -57
[本文引用: 1]
Liu Y K Tong H W Zhu Z W Chen L Zou J Zhang Y Q Jiao C H Gao B C Review on improvement of Fusarium head blight resistance in wheat
.J Triticeae Crops 2016 , 36 : 51 -57 (in Chinese with English abstract)
[本文引用: 1]
[12]
Brown-Guedira G Griffey C Kolb F McKendry A, Murphy J, Sanford D . Breeding FHB-resistant soft winter wheat: Progress and prospects
.Cereal Res Commun , 2008 , 36 : 31 -35
[本文引用: 2]
[13]
Bernardo A Bai G Yu J Kolb F Bockus W Dong Y Registration of near-isogenic winter wheat germplasm contrasting in Fhb1 for Fusarium head blight resistance
. J Plant Regist 2013 , 8 : 106 -108
[本文引用: 2]
[14]
Randhawa H S Asif M Pozniak C Clarke J M Graf R J Fox S L Humphreys D G Knox R E DePauw R M, Singh A K, Cuthbert R D, Hucl P, Spaner D . Application of molecular markers to wheat breeding in Canada
.Plant Breed , 2013 , 132 : 458 -471
[本文引用: 2]
[15]
Xie G Q Zhang M C Chakraborty S Liu C J The effect of 3BS locus of Sumai 3 on Fusarium head blight resistance in Australian wheats
.Aust J Exp Agric 2007 , 47 : 603 -607
[本文引用: 2]
[16]
Oda S. Advances in Wheat Genetics:From Genome to Field
. Berlin Heidelberg: Springer-Verlag Press , 2015 . pp 311 -318
[本文引用: 2]
[17]
张旭 , 任丽娟 , 谭秀云 , 藏宇辉 , 马鸿翔 , 秦浚川 , 陆维忠 . 利用分子标记研究小麦遗传群体的赤霉病抗性鉴定方法
. 南京大学学报(自然科学) , 2005 , 41 : 125 -132
[本文引用: 2]
Zhang X Ren L J Tan X Y Zang Y H Ma H X Qin J C Lu W Z Evaluation method of wheat Fusarium head blight resistance for genetic population based on molecular markers
.J Nanjing Univ Nat Sci ), 2005 , 41 : 125 -132 (in Chinese with English abstract)
[本文引用: 2]
[18]
中华人民共和国农业部
. 小麦抗病虫性评价技术规范——第4部分: 小麦抗赤霉病评价技术规范 , 2007 . pp 1 -6
[本文引用: 1]
Ministry of Agriculture of the People’s Republic of China
. Rules for Resistance Evaluation of Wheat to Diseases and Insect Pests—Part 4: Rules for Resistance Evaluation of Wheat to Wheat Scab (Fusarium graminearum Schwabe [Teleomorph Gibberella zeae (Schwein) Petch]) , 2007 . pp 1 -6 (in Chinese)
[本文引用: 1]
[19]
Rasheed A Wen W Gao F Zhai S Jin H Liu J Guo Q Zhang Y Dreisigacker S Xia X He Z Development and validation of KASP assays for genes underpinning key economic traits in bread wheat
.Theor Appl Genet , 2016 , 129 : 1843 -1860
[本文引用: 1]
[20]
刘秉华 , 杨丽 . “矮败”小麦的选育
. 中国农业科学 , 1990 , 23 (3 ): 86 -87
[本文引用: 1]
Liu B H Yang L Breeding of dwarfing and male-sterile wheat
.Sci Agric Sin 1990 , 23 (3 ): 86 -87 (in Chinese)
[本文引用: 1]
[21]
刘秉华 , 杨丽 . “矮败”小麦的选育及利用前景
. 科学通报 , 1991 , 36 : 306 -308
[本文引用: 1]
Liu B H Yang L Breeding and application of dwarfing and male-sterile wheat
. Chin Sci Bull , 1991 , 36 : 306 -308 (in Chinese)
[本文引用: 1]
[22]
陆维忠 . 小麦赤霉病抗性分子标记的筛选及其利用
. 江苏农业学报 , 2011 , 27 : 243 -249
[本文引用: 1]
Lu W Z Screening and application of molecular markers linked to wheat scab resistance
.Jiangsu J Agric Sci 2011 , 27 : 243 -249 (in Chinese with English abstract)
[本文引用: 1]
[23]
Liu S Pumphrey M Gill B Trick H Zhang J Dolezel J Chalhoub B Anderson J Toward positional cloning of Fhb1 , a major QTL for Fusarium head blight resistance in wheat
. Cereal Res Commun , 2008 , 36 : 195 -201
[本文引用: 1]
[24]
Zhou W C Kolb F L Bai G H Domier L L Yao J B Effect of individual Sumai 3 chromosomes on resistance to scab spread within spikes and deoxynivalenol accumulation within kernels in wheat
. Hereditas , 2002 , 137 : 81 -89
[本文引用: 1]
[25]
Pumphrey M O Bernardo R Anderson J A Validating the Fhb1 QTL for Fusarium head blight resistance in near-isogenic wheat lines developed from breeding populations
. Crop Sci , 2007 , 47 : 200 -206
[本文引用: 1]
[26]
陆维忠 , 马鸿翔 . 高产抗赤霉病小麦新品种生选6号的选育
. 江苏农业科学 , 2010 , 38 (3 ): 153 -154
[本文引用: 1]
Lu W Z Ma H X Breeding of wheat cultivar Shengxuan 6 for high-resistant Fusarium head blight
.Jiangsu Agric Sci 2010 , 38 (3 ): 153 -154 (in Chinese)
[本文引用: 1]
[27]
姚金保 , 陆维忠 . 小麦新品种生选6号的丰产稳产性和赤霉病抗性分析
. 麦类作物学报 , 2012 , 32 : 793 -798
[本文引用: 1]
Yao J B Lu W Z Analysis of yield potential, yield stability and resistance to Fusarium head blight of new wheat cultivar Shengxuan 6
.J Triticeae Crops 2012 , 32 : 793 -798 (in Chinese with English abstract)
[本文引用: 1]
[28]
张旭 , 姜朋 , 叶人元 , 吴磊 , 张瑜 , 马鸿翔 . 宁麦9号及其衍生品种的赤霉病抗性分析与抗性溯源
. 分子植物育种 , 2017 , 15 : 1053 -1060
[本文引用: 1]
Zhang X Jiang P Ye R Y Wu L Zhang Y Ma H X Evaluation and source tracing of resistance to Fusarium head blight in wheat variety Ningmai 9 and its derivatives
.Mol Plant Breed 2017 , 15 : 1053 -1060 (in Chinese with English abstract)
[本文引用: 1]
Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F
1
2009
... 小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1 ] .受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2 ] .在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2 ] .然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
中国小麦赤霉病的危害及抗性遗传改良
2
2012
... 小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1 ] .受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2 ] .在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2 ] .然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
... [2 ].然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
中国小麦赤霉病的危害及抗性遗传改良
2
2012
... 小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1 ] .受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2 ] .在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2 ] .然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
... [2 ].然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
气温变化背景下中国黄淮旱地冬小麦农艺性状的变化特征——以山西临汾为例
1
2015
... 小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1 ] .受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2 ] .在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2 ] .然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
气温变化背景下中国黄淮旱地冬小麦农艺性状的变化特征——以山西临汾为例
1
2015
... 小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1 ] .受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2 ] .在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2 ] .然而, 作为我国小麦主产区的黄淮冬麦区[3 ] , 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病.发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一. ...
DNA markers for Fusarium head blight resistance QTLs in two wheat populations
3
2001
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 抗赤霉病基因Fhb1 位于3B染色体Xgwm533.1 - Xgwm493 标记区间内, 抗性等位变异来自苏麦3号[4 ,5 ] .陆维忠[22 ] 采用4个赤霉病抗病品种与2个感病品种构建的6个群体进行抗赤霉病QTL发掘证实, 抗病品种携带的主效QTL均在3B染色体Xgwm533.1 - Xgwm493 之间, 单个标记的选择效率达到70%.在此基础上, Liu等[23 ] 进一步将Fhb1 定位在231 kb区间, 并开发了共显性特异标记UMN10, 其选择效率明显高于SSR标记.在本研究中, 利用Fhb1 的功能KASP标记鉴定发现, 含有Fhb1 基因的品种一般对赤霉病的抗性较好, 例如, 中抗赤霉病品种(系)生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798均含有Fhb1 基因; 同时也发现有些赤霉病抗性较好的品种不含有Fhb1 基因(例如扬麦158), 但未发现既具有该标记又高感赤霉病的品种.这一方面说明Fhb1 基因对赤霉病抗性的有效性, 另一方面也说明除Fhb1 基因外, 有些品种可能携带其他抗赤霉病基因. ...
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
Fine mapping Fhb1 , a major gene controlling Fusarium head blight resistance in bread wheat(Triticum aestivum L.)
2
2006
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 抗赤霉病基因Fhb1 位于3B染色体Xgwm533.1 - Xgwm493 标记区间内, 抗性等位变异来自苏麦3号[4 ,5 ] .陆维忠[22 ] 采用4个赤霉病抗病品种与2个感病品种构建的6个群体进行抗赤霉病QTL发掘证实, 抗病品种携带的主效QTL均在3B染色体Xgwm533.1 - Xgwm493 之间, 单个标记的选择效率达到70%.在此基础上, Liu等[23 ] 进一步将Fhb1 定位在231 kb区间, 并开发了共显性特异标记UMN10, 其选择效率明显高于SSR标记.在本研究中, 利用Fhb1 的功能KASP标记鉴定发现, 含有Fhb1 基因的品种一般对赤霉病的抗性较好, 例如, 中抗赤霉病品种(系)生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798均含有Fhb1 基因; 同时也发现有些赤霉病抗性较好的品种不含有Fhb1 基因(例如扬麦158), 但未发现既具有该标记又高感赤霉病的品种.这一方面说明Fhb1 基因对赤霉病抗性的有效性, 另一方面也说明除Fhb1 基因外, 有些品种可能携带其他抗赤霉病基因. ...
Mapping of Fhb2 on chromosome 6BS: a gene controlling Fusarium head blight field resistance in bread wheat(Triticum aestivum L.)
1
2007
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
Molecular cytogenetic characterization of alien introgressions with gene Fhb3 for resistance to Fusarium head blight disease of wheat
1
2008
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
High-density mapping of the major FHB resistance geneFhb7 derived from Thinopyrum ponticum and its pyramiding with Fhb1 by marker-assisted selection
1
2015
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
Fine mapping Fhb4 , a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.)
1
2010
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
Precise mapping Fhb5 , a major QTL conditioning resistance to Fusarium infection in bread wheat ( Triticum aestivum L.)
1
2011
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
小麦赤霉病抗性改良研究进展
1
2016
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
小麦赤霉病抗性改良研究进展
1
2016
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
Breeding FHB-resistant soft winter wheat: Progress and prospects
2
2008
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
Registration of near-isogenic winter wheat germplasm contrasting in Fhb1 for Fusarium head blight resistance
2
2013
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
Application of molecular markers to wheat breeding in Canada
2
2013
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
The effect of 3BS locus of Sumai 3 on Fusarium head blight resistance in Australian wheats
2
2007
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
Advances in Wheat Genetics:From Genome to Field
2
2015
... 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4 ,5 ] 、Fhb2 [6 ] 、Fhb3 [7 ] 和Fhb7 [8 ] , 以及抗侵入(Type I)相关基因, 包括Fhb4 [9 ] 和Fhb5 [10 ] .其中, 位于3B染色体上的Fhb1 是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11 ] .利用Fhb1 分子标记辅助选择技术, 已在美国[12 ,13 ] 、加拿大[14 ] 、澳大利亚[15 ] 、日本[16 ] 等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高.黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定.稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程.本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1 基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1 基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1 基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持. ...
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
利用分子标记研究小麦遗传群体的赤霉病抗性鉴定方法
2
2005
... 2016年10月, 将来自上述6个回交群体携带Fhb1 基因的11个可育株和11个不携带Fhb1 基因的可育株、轮回亲本周麦16、中感赤霉病对照品种淮麦20、中抗赤霉病对照品种扬麦158和高感赤霉病的对照品种安农8455种植在江苏省农业科学院(南京市)赤霉病鉴定圃, 采用单花滴注法[17 ] 人工接种鉴定.采用完全随机区组设计, 单行区, 行长2.0 m, 行距0.3 m, 株距5.0 cm, 2次重复.单花滴注的赤霉菌孢子液10 μL, 孢子浓度为5 × 105 mL-1 .在始花期, 每个株系接种10穗, 套袋保湿72 h后, 在弥雾保湿条件下继续生长, 接种后21 d调查病小穗数, 作为赤霉病抗性评价指标[17 ] . ...
... [17 ]. ...
利用分子标记研究小麦遗传群体的赤霉病抗性鉴定方法
2
2005
... 2016年10月, 将来自上述6个回交群体携带Fhb1 基因的11个可育株和11个不携带Fhb1 基因的可育株、轮回亲本周麦16、中感赤霉病对照品种淮麦20、中抗赤霉病对照品种扬麦158和高感赤霉病的对照品种安农8455种植在江苏省农业科学院(南京市)赤霉病鉴定圃, 采用单花滴注法[17 ] 人工接种鉴定.采用完全随机区组设计, 单行区, 行长2.0 m, 行距0.3 m, 株距5.0 cm, 2次重复.单花滴注的赤霉菌孢子液10 μL, 孢子浓度为5 × 105 mL-1 .在始花期, 每个株系接种10穗, 套袋保湿72 h后, 在弥雾保湿条件下继续生长, 接种后21 d调查病小穗数, 作为赤霉病抗性评价指标[17 ] . ...
... [17 ]. ...
中华人民共和国农业部
1
2007
... 2016年11月, 将上述用于人工接种鉴定的所有可育株、轮回亲本、中感对照品种淮麦20种植在福建省南平市农业科学研究所赤霉病田间鉴定圃进行自然鉴定.采用完全随机区组设计, 单行区, 行长1.0 m, 行距0.3 m, 株距5.0 cm, 3次重复.扬花后20 d, 每个材料每重复调查15穗.采用病情指数作为田间病圃鉴定的评价指标[18 ] . ...
中华人民共和国农业部
1
2007
... 2016年11月, 将上述用于人工接种鉴定的所有可育株、轮回亲本、中感对照品种淮麦20种植在福建省南平市农业科学研究所赤霉病田间鉴定圃进行自然鉴定.采用完全随机区组设计, 单行区, 行长1.0 m, 行距0.3 m, 株距5.0 cm, 3次重复.扬花后20 d, 每个材料每重复调查15穗.采用病情指数作为田间病圃鉴定的评价指标[18 ] . ...
Development and validation of KASP assays for genes underpinning key economic traits in bread wheat
1
2016
... 采用降落PCR扩增, 程序为95°C热启动15 min, 9个降落循环(95°C 20 s, 65°C 20 s, 从65°C每个循环降低1°C), 95°C变性10 s, 57°C退火60 s, 35个循环, 延伸5 min [19 ] .反应结束后, 在酶标仪SYNERGY H1MF microplate reader (BioTek Instruments, Inc., 美国)上读取荧光数据, 并导入Kluster Caller software version 2.24.0.11 (http://kbioscience. co.uk/)作基因分型. ...
“矮败”小麦的选育
1
1990
... 矮败小麦具有雄性败育彻底、不育性稳定、异交结实率高的优点, 同时, 由于显性矮秆基因Rht-D1c 与显性雄性不育基因Ms2 紧密连锁, 矮秆不育株与正常株高的可育株很容易区别, 简化了可育株和不育株的人工鉴定和标记过程, 是理想的小麦遗传改良工具[20 ,21 ] .我们把矮败小麦的Rht-D1c 和Ms2 基因回交转育到黄淮冬麦区推广品种和骨干亲本, 培育出了这些品种和骨干亲本的近等基因系, 例如, 矮败周麦16、矮败郑麦9023、矮败济麦22、矮败轮选987等.本研究利用矮败周麦16与具有Fhb1 基因的品种(系)杂交和回交, 利用KASP标记对后代进行Fhb1 基因型分析, 对携带Fhb1 基因的矮秆不育株, 继续与黄淮麦区的品种(系)回交; 对携带Fhb1 基因的可育株进行赤霉病抗性鉴定和农艺性状选择, 培育符合黄淮冬麦区生态类型、赤霉病抗性得到显著改良的新品种.这种策略发挥了矮败小麦杂交方便、回交后代群体大的优势, 有利于较快地选择出既具有轮回亲本优良的农艺性状、赤霉病抗性又得到改良的新品种. ...
“矮败”小麦的选育
1
1990
... 矮败小麦具有雄性败育彻底、不育性稳定、异交结实率高的优点, 同时, 由于显性矮秆基因Rht-D1c 与显性雄性不育基因Ms2 紧密连锁, 矮秆不育株与正常株高的可育株很容易区别, 简化了可育株和不育株的人工鉴定和标记过程, 是理想的小麦遗传改良工具[20 ,21 ] .我们把矮败小麦的Rht-D1c 和Ms2 基因回交转育到黄淮冬麦区推广品种和骨干亲本, 培育出了这些品种和骨干亲本的近等基因系, 例如, 矮败周麦16、矮败郑麦9023、矮败济麦22、矮败轮选987等.本研究利用矮败周麦16与具有Fhb1 基因的品种(系)杂交和回交, 利用KASP标记对后代进行Fhb1 基因型分析, 对携带Fhb1 基因的矮秆不育株, 继续与黄淮麦区的品种(系)回交; 对携带Fhb1 基因的可育株进行赤霉病抗性鉴定和农艺性状选择, 培育符合黄淮冬麦区生态类型、赤霉病抗性得到显著改良的新品种.这种策略发挥了矮败小麦杂交方便、回交后代群体大的优势, 有利于较快地选择出既具有轮回亲本优良的农艺性状、赤霉病抗性又得到改良的新品种. ...
“矮败”小麦的选育及利用前景
1
1991
... 矮败小麦具有雄性败育彻底、不育性稳定、异交结实率高的优点, 同时, 由于显性矮秆基因Rht-D1c 与显性雄性不育基因Ms2 紧密连锁, 矮秆不育株与正常株高的可育株很容易区别, 简化了可育株和不育株的人工鉴定和标记过程, 是理想的小麦遗传改良工具[20 ,21 ] .我们把矮败小麦的Rht-D1c 和Ms2 基因回交转育到黄淮冬麦区推广品种和骨干亲本, 培育出了这些品种和骨干亲本的近等基因系, 例如, 矮败周麦16、矮败郑麦9023、矮败济麦22、矮败轮选987等.本研究利用矮败周麦16与具有Fhb1 基因的品种(系)杂交和回交, 利用KASP标记对后代进行Fhb1 基因型分析, 对携带Fhb1 基因的矮秆不育株, 继续与黄淮麦区的品种(系)回交; 对携带Fhb1 基因的可育株进行赤霉病抗性鉴定和农艺性状选择, 培育符合黄淮冬麦区生态类型、赤霉病抗性得到显著改良的新品种.这种策略发挥了矮败小麦杂交方便、回交后代群体大的优势, 有利于较快地选择出既具有轮回亲本优良的农艺性状、赤霉病抗性又得到改良的新品种. ...
“矮败”小麦的选育及利用前景
1
1991
... 矮败小麦具有雄性败育彻底、不育性稳定、异交结实率高的优点, 同时, 由于显性矮秆基因Rht-D1c 与显性雄性不育基因Ms2 紧密连锁, 矮秆不育株与正常株高的可育株很容易区别, 简化了可育株和不育株的人工鉴定和标记过程, 是理想的小麦遗传改良工具[20 ,21 ] .我们把矮败小麦的Rht-D1c 和Ms2 基因回交转育到黄淮冬麦区推广品种和骨干亲本, 培育出了这些品种和骨干亲本的近等基因系, 例如, 矮败周麦16、矮败郑麦9023、矮败济麦22、矮败轮选987等.本研究利用矮败周麦16与具有Fhb1 基因的品种(系)杂交和回交, 利用KASP标记对后代进行Fhb1 基因型分析, 对携带Fhb1 基因的矮秆不育株, 继续与黄淮麦区的品种(系)回交; 对携带Fhb1 基因的可育株进行赤霉病抗性鉴定和农艺性状选择, 培育符合黄淮冬麦区生态类型、赤霉病抗性得到显著改良的新品种.这种策略发挥了矮败小麦杂交方便、回交后代群体大的优势, 有利于较快地选择出既具有轮回亲本优良的农艺性状、赤霉病抗性又得到改良的新品种. ...
小麦赤霉病抗性分子标记的筛选及其利用
1
2011
... 抗赤霉病基因Fhb1 位于3B染色体Xgwm533.1 - Xgwm493 标记区间内, 抗性等位变异来自苏麦3号[4 ,5 ] .陆维忠[22 ] 采用4个赤霉病抗病品种与2个感病品种构建的6个群体进行抗赤霉病QTL发掘证实, 抗病品种携带的主效QTL均在3B染色体Xgwm533.1 - Xgwm493 之间, 单个标记的选择效率达到70%.在此基础上, Liu等[23 ] 进一步将Fhb1 定位在231 kb区间, 并开发了共显性特异标记UMN10, 其选择效率明显高于SSR标记.在本研究中, 利用Fhb1 的功能KASP标记鉴定发现, 含有Fhb1 基因的品种一般对赤霉病的抗性较好, 例如, 中抗赤霉病品种(系)生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798均含有Fhb1 基因; 同时也发现有些赤霉病抗性较好的品种不含有Fhb1 基因(例如扬麦158), 但未发现既具有该标记又高感赤霉病的品种.这一方面说明Fhb1 基因对赤霉病抗性的有效性, 另一方面也说明除Fhb1 基因外, 有些品种可能携带其他抗赤霉病基因. ...
小麦赤霉病抗性分子标记的筛选及其利用
1
2011
... 抗赤霉病基因Fhb1 位于3B染色体Xgwm533.1 - Xgwm493 标记区间内, 抗性等位变异来自苏麦3号[4 ,5 ] .陆维忠[22 ] 采用4个赤霉病抗病品种与2个感病品种构建的6个群体进行抗赤霉病QTL发掘证实, 抗病品种携带的主效QTL均在3B染色体Xgwm533.1 - Xgwm493 之间, 单个标记的选择效率达到70%.在此基础上, Liu等[23 ] 进一步将Fhb1 定位在231 kb区间, 并开发了共显性特异标记UMN10, 其选择效率明显高于SSR标记.在本研究中, 利用Fhb1 的功能KASP标记鉴定发现, 含有Fhb1 基因的品种一般对赤霉病的抗性较好, 例如, 中抗赤霉病品种(系)生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798均含有Fhb1 基因; 同时也发现有些赤霉病抗性较好的品种不含有Fhb1 基因(例如扬麦158), 但未发现既具有该标记又高感赤霉病的品种.这一方面说明Fhb1 基因对赤霉病抗性的有效性, 另一方面也说明除Fhb1 基因外, 有些品种可能携带其他抗赤霉病基因. ...
Toward positional cloning of Fhb1 , a major QTL for Fusarium head blight resistance in wheat
1
2008
... 抗赤霉病基因Fhb1 位于3B染色体Xgwm533.1 - Xgwm493 标记区间内, 抗性等位变异来自苏麦3号[4 ,5 ] .陆维忠[22 ] 采用4个赤霉病抗病品种与2个感病品种构建的6个群体进行抗赤霉病QTL发掘证实, 抗病品种携带的主效QTL均在3B染色体Xgwm533.1 - Xgwm493 之间, 单个标记的选择效率达到70%.在此基础上, Liu等[23 ] 进一步将Fhb1 定位在231 kb区间, 并开发了共显性特异标记UMN10, 其选择效率明显高于SSR标记.在本研究中, 利用Fhb1 的功能KASP标记鉴定发现, 含有Fhb1 基因的品种一般对赤霉病的抗性较好, 例如, 中抗赤霉病品种(系)生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798均含有Fhb1 基因; 同时也发现有些赤霉病抗性较好的品种不含有Fhb1 基因(例如扬麦158), 但未发现既具有该标记又高感赤霉病的品种.这一方面说明Fhb1 基因对赤霉病抗性的有效性, 另一方面也说明除Fhb1 基因外, 有些品种可能携带其他抗赤霉病基因. ...
Effect of individual Sumai 3 chromosomes on resistance to scab spread within spikes and deoxynivalenol accumulation within kernels in wheat
1
2002
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
Validating the Fhb1 QTL for Fusarium head blight resistance in near-isogenic wheat lines developed from breeding populations
1
2007
... 在抗病基因作图群体中, Fhb1 能够在不同的遗传背景下解释20%~60%的表型变异[4 ,24 -25 ] , 是公认的用于赤霉病改良的优异基因.目前, 该基因也是仅有几个可以用于小麦分子标记辅助育种的目标基因之一, 并取得一定的效果[12 ,13 ,14 ,15 ,16 ] .与轮回亲本周麦16相比, 本研究6个回交群体中携带供体Fhb1 基因的株系平均病小穗数减少, 平均病情指数显著降低, 抗性水平达到或者超过中感赤霉病的对照品种淮麦20, 而且所有回交后代平均病小穗数显著低于不含有Fhb1 的回交后代, 表明利用Fhb1 基因分子标记辅助选择技术能够有效地提高该麦区小麦品种赤霉病抗性水平.与赤霉病抗性有关的基因很多, 育种上要在短时间内把多个抗赤霉病基因聚合在一起难度很大, 黄淮麦区抗赤霉病育种可以采取循序渐进的办法, 通过大群体回交结合分子标记辅助选择, 快速把Fhb1 基因转育到主栽品种或苗头品系中, 使育成品种对赤霉病的抗性水平由目前的高感变为中感, 然后再进行多个抗赤霉病基因的聚合, 进一步提高品种对赤霉病的抗性水平. ...
高产抗赤霉病小麦新品种生选6号的选育
1
2010
... 尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异.田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强.生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当.分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] .本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究.宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] .与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关. ...
高产抗赤霉病小麦新品种生选6号的选育
1
2010
... 尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异.田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强.生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当.分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] .本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究.宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] .与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关. ...
小麦新品种生选6号的丰产稳产性和赤霉病抗性分析
1
2012
... 尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异.田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强.生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当.分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] .本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究.宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] .与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关. ...
小麦新品种生选6号的丰产稳产性和赤霉病抗性分析
1
2012
... 尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异.田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强.生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当.分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] .本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究.宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] .与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关. ...
宁麦9号及其衍生品种的赤霉病抗性分析与抗性溯源
1
2017
... 尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异.田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强.生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当.分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] .本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究.宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] .与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关. ...
宁麦9号及其衍生品种的赤霉病抗性分析与抗性溯源
1
2017
... 尽管不同供体携带相同的Fhb1 基因, 但回交后代的赤霉病抗性存在显著差异.田间病圃自然发病和单花滴注两种鉴定方法的结果表明, 生选6号回交后代比其他供体的后代对赤霉病的抗性更强.生选6号是以中感赤霉病的宁麦8号为母本与中抗赤霉病的宁麦9号为父本杂交选育而成[26 ] , 具有很好的丰产性, 而且在赤霉病抗性上表现超亲遗传, 抗性水平达到高抗, 与苏麦3号相当.分子标记检测发现生选6号具有更多其双亲不具备但在苏麦3号和望水白中存在的抗性基因座[27 ] .本研究中生选6号回交后代表现中抗的赤霉病抗性是否与其具有其他抗性基因座有关需要进一步研究.宁麦9号中抗赤霉病, 它的衍生品种宁麦13具有更多的赤霉病抗性基因座, 而且在赤霉病抗性上优于宁麦9号[28 ] .与其亲本不同, 宁麦9号的回交后代在抗病性上优于宁麦13的回交后代, 这可能与遗传背景效应有关. ...
 , 宿振起
, 宿振起


{kind=link}
{kind=link}
{kind=link}
{kind=link}