紫苏二酰基甘油酰基转移酶2基因克隆与功能研究
鲁庚,唐鑫,陆俊杏,李丹,胡秋芸,胡田,张涛
作物学报
2020, 46 ( 8):
1283-1290.
DOI: 10.3724/SP.J.1006.2020.94192
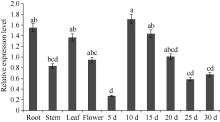
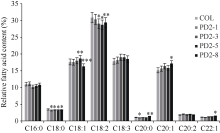
二酰基甘油酰基转移酶(diacylglycerol acyltransferase, DGAT)是植物合成三酰甘油(TAG)最后一步的关键酶, 其中DGAT2在某些植物的种子油中能选择性积累更多不饱和脂肪酸。本文成功克隆了紫苏二酰基甘油酰基转移酶2基因(PfDGAT2), 并进行生物信息学分析。PfDGAT2实时荧光定量结果表明, 不同器官中PfDGAT2基因均有表达, 10 d种子的表达量最高, 在根中的表达量次之,在种子发育中后期, PfDGAT2表达量逐渐降低。与野生型拟南芥相比, 过表达PfDGAT2拟南芥种子含油率提高了21.68%~77.89%, 其中种子含油率增加最多的4个株系, 其亚麻酸(C18:3)增加4.57%, 花生一烯酸(C20:1)增加7.44%, 花生二烯酸(C20:2)增加5.4%, 二十二一烯酸(C22:1)增加10.37%, 而棕榈酸(C16:0)、硬脂酸(C18:0)和亚油酸(C18:2)含量分别降低了3.47%、6.64%和4.83%, 油酸(C18:1)和花生酸(C20:0)分别只降低了0.18%和1.91%。本研究结果表明, 紫苏PfDGAT2基因不仅能提高种子含油率, 还能促进亚麻酸、花生一烯酸等不饱和脂肪酸的积累, 这为研究植物不饱和脂肪酸的合成积累提供了参考及理论依据。
引物名称
Primer name | 引物序列
Sequences (5'-3') | | PfDGAT2 F | CTCGCTTACTGCTACTTCAATG | | PfDGAT2 R | CGACAATTAGAGAATCCTGAGC | | PfDGAT2-DL F | AGTCCGAGCCCAACGGCGATGTCAG | | PfDGAT2-DL R | GGATGCCCAACCCTTGCTTTGTGCC | | Actin F | AGACCTTCAATGTGCCAGCCA | | Actin R | CACGACCAGCAAGATCCAACC | | 18S RNAF | CGGCGACGCATCATTCAAA | | DGAT2-BglII F | ACTCTTGACCATGGTAGATCTGGAGTCCGAGCCCAACGGCG | | DGAT2-BglII R | GGACGTAAACTAGTCAGATCTAGAATCCTGAGCTCTAAGTCG | | JD-F | TTTCATTTGGAGAGAACACGGGGGA | | JD-R | CGCTGATCAATTCCACAGTTTTCGC |
View table in article
表1
PCR引物
正文中引用本图/表的段落
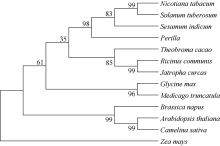
TAG的生物合成涉及质体中脂肪酸的生物合成和内质网中TAG的组装2个主要阶段。脂肪酸的生物合成主要是乙酰辅酶A羧化酶(ACCase)将乙酰辅酶A活化为丙二酰辅酶A, 然后将丙二酰基从CoA转移到酰基载体蛋白(ACP)作为酰基链载体, 合成的脂肪酸则输出到内质网(ER)[6]。在肯尼迪途径中, 三磷酸甘油酰基转移酶(GPAT)、溶血磷脂酰基转移酶(LPAAT)和二酰基甘油酰基转移酶(DGAT)催化三磷酸甘油骨架链上酰化反应, 最后生成三酰甘油[7,8]。其中DGAT是用于TAG合成的最后一种酶, 它将第3个酰基链添加到DAG并产生TAG。目前, 在植物中鉴定出DGAT1、DGAT2、DGAT3[8,9]和WS/ DGAT[10]4种类型的DGAT。DGAT1和DGAT2在真核生物中普遍存在, 它们是2个结构不同的膜结合酰基转移酶, 没有序列同源性[12]。另外, DGAT1和DGAT2在底物特异性和表达模式等方面也不同[12,13,14]。Saha等[8]在花生子叶中克隆出了第3类可溶性DGAT酶, 命名为DGAT3, 定位于细胞质中。第4种类型的DGAT酶是从醋酸钙不动杆菌ADP1鉴定的双功能膜相关酶, 同时具有WS和DGAT活性[15,16]。在拟南芥和矮牵牛中的WS/DGAT同源物主要是催化蜡酯的合成[16,17]。有研究表明, 拟南芥的二酰基甘油酰基转移酶1 (DGAT1)是油脂中正常TAG积累所必需的[18,19,20]。但是, 拟南芥中的AtDGAT2作用尚不清楚, 因为dgat1突变体与AtDGAT2敲除突变体杂交后种子的含油量并没有明显降低[12]。与拟南芥相比, 蓖麻胚乳中的DGAT2在种子中积累了大量的TAG (含90%以上蓖麻酸)[21]。此外, Shockey等[13]表明, DGAT2可能参与异常脂肪酸、桐油酸和桐油的选择性积累。随后Oelkers[22]表明, 蓖麻DGAT2能与蓖麻羟化酶共同表达, 在拟南芥种子中使羟基脂肪酸积累加倍(从~17%到~30%)。Li等[23]也证明了斑鸠菊、大戟和琉璃菊中的PDAT和DGAT2积累了更高的环氧和羟基脂肪酸。Yuan等[24]通过在拟南芥中过表达油棕EgDGAT2, 提高了拟南芥种子TAG中多不饱和脂肪酸亚油酸和亚麻酸的含量, 硬脂酸和花生酸的比例也相应降低。在最近的研究中, Zheng等[25]在拟南芥中过表达椰子CoDGAT2, 导致拟南芥种子的亚油酸含量显著增加, 二十碳一烯酸和花生酸的比例降低。因此, DGAT2在某些植物种子油中选择性积累不饱和脂肪酸方面所起的作用比DGAT1更为显著[12]。
1.1.1 植物材料 紫苏、拟南芥种子由重庆师范大学油用牡丹种质资源创新与利用重点实验室保存。于2018年4月将紫苏种植于重庆师范大学植物园, 9月采集花、叶、茎、根以及花后5、10、15、20、25、30 d不同发育阶段的种子, 液氮冷冻运输, 然后在-80℃冰箱保存待用。所有拟南芥种子均经2%次氯酸钠表面消毒, 播种在1/2 MS培养基上, 将22℃长日条件下的2周龄植物移栽到土壤(营养土:细土:珍珠岩 = 1.0:3.0:0.5)中。
1.2.2 PfDGAT2基因的克隆 从实验室前期紫苏转录组测序数据中挑选出功能注释为DGAT2的基因[29], 以拟南芥AtDGAT2为参照, 利用BioEdit软件比对分析, 获得紫苏PfDGAT2全长。采用Primer Premier 5.0软件分别设计5'端及3'端全长特异性引物。反应引物为PfDGAT2 F和PfDGAT2 R (表1)。根据设计的引物以反转录的cDNA为模板进行PCR扩增, 将扩增产物纯化后与载体pMD-T 19连接, 转化大肠杆菌DH5α感受态, 筛选蓝白斑阳性克隆, PCR鉴定挑选单克隆送测序, 引物及测序均由英潍捷基有限公司完成。
1.2.4 PfDGAT2基因组织特异性表达分析 根据已克隆PfDGAT2的cDNA序列分析结果, 设计荧光定量PCR特异引物(表1), 以紫苏β-actin和18S RNA作为内参基因, 参照试剂盒SYBR Premix Ex Taq II在CFX96 Real-Time PCR Detection System仪器(Bio-Rad公司)上进行荧光定量PCR检测, 反应体系为10 μL, 包含SYBR Green supermix 5.0 μL、上下游引物各0.5 μL、Template 4.0 μL; 反应条件为95℃预变性60 s; 95℃变性10 s, 60℃退火20 s, 40个循环; 反应结束后再升温至95℃, 最后进行溶解曲线分析。每组实验设置3次生物学重复和3次技术重复。
1.2.5 植物表达载体构建及转化 用限制性内切酶Bgl II酶切植物表达载体pCAMBIA1303, 设计带有Bgl II酶切位点的1对引物DGAT2-BglII F和DGAT2-BglII R (表1), 采用In-Fusion无缝克隆得到过表达重组质粒 WT: Pro35SPfDGAT2, 将重组质粒转化到农杆菌中, 利用农杆菌介导的花序侵染法转化野生型拟南芥Col-0。农杆菌侵染的当代植株为T0代, 单株收的每代种子经1/2 MS培养基(含30 mg mL-1潮霉素)筛选后移栽至温室生长, 鉴定阳性苗使用分子鉴定, 分子鉴定引物为JD-F、JD-R (表1)。
本文的其它图/表
|