欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (12): 3154-3161.doi: 10.3724/SP.J.1006.2023.31009
刘小娟1,2( ), 柳欣2, 张明虎2, 郝明2, 甯顺腙2, 袁中伟2, 黄林2, 刘登才1,2, 张连全1,2,*()
), 柳欣2, 张明虎2, 郝明2, 甯顺腙2, 袁中伟2, 黄林2, 刘登才1,2, 张连全1,2,*()
LIU Xiao-Juan1,2(), LIU Xin2, ZHANG Ming-Hu2, HAO Ming2, NING Shun-Zong2, YUAN Zhong-Wei2, HUANG Lin2, LIU Deng-Cai1,2, ZHANG Lian-Quan1,2,*()
摘要:
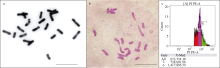
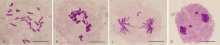
未减数配子的结合实现染色体自动加倍, 是多倍体物种起源的重要途径, 也是提高作物单倍体育种效率的重要手段。我们前期从四倍体小麦发掘出控制未减数配子形成的强效QTL位点QTug.sau-3B, 并通过人工合成小麦为“桥梁”, 将其导入到综合农艺性状优良的小麦新品系中。本实验使用5份含未减数配子基因的优良小麦新品系与不含未减数配子基因的小麦推广品种的F1杂种作母本与3份白茅(Imperata cylindrica)进行远缘杂交, 共授粉4610朵小花, 结实1965粒, 经幼胚拯救获得244个幼胚, 其中50个幼胚发育正常生长为50个小麦单倍体植株。由于小麦单倍体植株未减数配子基因的表达易受环境影响, 因此, 对单倍体植株在相同光周期(18 h光照/6 h黑暗)下进行了不同温度25℃/18℃、25℃/15℃和25℃/10℃处理, 结果表明, 25℃/18℃和25℃/10℃条件下编号为H31单倍体植株能够结实, 自交结实率分别为4.35%和2.41%。该研究结果为建立“基于小麦-白茅杂交实现染色体消除和未减数配子基因实现染色体自动加倍”的小麦单倍体育种技术提供了参考。
| [1] | Raina S K. Doubled haploid breeding in cereals. Plant Breed Rev, 1997, 15: 141-186. |
| [2] |
Machii H, Mizuno H, Hirabayashi T, Li H, Hagio T. Screening wheat genotypes for high callus induction and regeneration capability from anther and immature embryo cultures. Plant Cell Tissue Organ Cult, 1998, 53: 67-74.
doi: 10.1023/A:1006023725640 |
| [3] |
Cistué L, Soriano M, Castillo A M, Vallés M P, Sanz J M, Echávarri B. Production of doubled haploids in durum wheat (Triticum turgidum L.) through isolated microspore culture. Plant Cell Rep, 2006, 25: 257-264.
pmid: 16220343 |
| [4] | Touraev A, Pfosser M, Heberle-Bors E. The microspore: a haploid multipurpose cell. Adv Bot Res, 2001, 35: 53-109. |
| [5] | Zhu H, Li C, Gao C. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nat Rew Mol Cell Biol, 2020, 21: 661-677. |
| [6] |
Liu H Y, Wang K, Jia Z M, Gong Q, Lin Z S, Du L P, Pei X W, Ye X G. Efficient induction of haploid plants in wheat by editing of TaMTL using an optimized Agrobacterium-mediated CRISPR system. J Exp Bot, 2020, 71: 1337-1349.
doi: 10.1093/jxb/erz529 |
| [7] |
Liu C X, Zhong Y, Qi X L, Chen M, Liu Z K, Chen C, Tian X L, Li J L, Jiao Y Y, Wang D, Wang Y W, Li M R, Xin M M, Liu W X, Jin W W, Chen S J. Extension of the in vivo haploid induction system from diploid maize to hexaploid wheat. Plant Biotechnol J, 2020, 18: 316-318.
doi: 10.1111/pbi.v18.2 |
| [8] |
Barclay I R. High frequencies of haploid production in wheat (Triticum aestivum) by chromosome elimination. Nature, 1975, 256: 410-411.
doi: 10.1038/256410a0 |
| [9] |
Zenkteler M, Nitzsche W. Wide hybridization experiments in cereals. Theor Appl Genet, 1984, 68: 311-315.
doi: 10.1007/BF00267883 pmid: 24257639 |
| [10] |
Inagaki M, Mujeeb-Kazi A. Comparison of polyhaploid production prequencies in crosses of hexaploid wheat with maize, pearl millet and sorghum. Jpn J Breed, 1995, 45: 157-161.
doi: 10.1270/jsbbs1951.45.157 |
| [11] |
Riera-Lizarazu O, Mujeeb-Kazi A. Polyhaploid production in the Triticeae: wheat × Tripsacum crosses. Crop Sci, 1993, 33: 973-976.
doi: 10.2135/cropsci1993.0011183X003300050020x |
| [12] |
Mochida K. Production of wheat doubled haploids by pollination with Job’s Tears (Coix lachryma-jobi L.). J Hered, 2001, 92: 81-83.
doi: 10.1093/jhered/92.1.81 pmid: 11336235 |
| [13] |
Chaudhary H K, Sethi G S, Singh S, Pratap A, Sharma S. Efficient haploid induction in wheat by using pollen of Imperata cylindrica. Plant Breed, 2005, 124: 96-98.
doi: 10.1111/pbr.2005.124.issue-1 |
| [14] | Liu D C, Zhang H G, Zhang L Q, Yuan Z W, Hao M, Zheng Y L. Distant hybridization: a tool for interspecific manipulation of chromosomes. In: Pratap A, Kumar J, eds. Alien Gene Transfer in Crop Plants, Volume 1: Innovations, Methods and Risk Assessment. New York: Springer, 2014. pp 25-42. |
| [15] | Chaudhary H K, Kaila V, Rather S A, Tayeng T. Distant hybridization and doubled-haploidy breeding. In: Pratap A, Kumar J, eds. Alien Gene Transfer in Crop Plants, Volume 1: Innovations, Methods and Risk Assessment. New York: Springer, 2014. pp 143-164. |
| [16] |
Chan S W L. Chromosome engineering: power tools for plant genetics. Trends Biotech, 2010, 28: 605-610.
doi: 10.1016/j.tibtech.2010.09.002 |
| [17] | Cheng K T, Chou C H. Ecotypic variation of Imperata cylindrica populations in Taiwan: I. Morphological and molecular evidences. Bot Stud, 1997, 38: 215-223. |
| [18] | Chaudhary H K. Dynamics of wheat × Imperata cylindrica: a new chromosome elimination mediated system for efficient haploid induction in wheat. In: Appels R, Eastwood R, Lagudah E, Langridge P, Mackay M, Mcintyre L, Sharp P, eds. Proceedings of the 11th International Wheat Genetics Symposium. Sydney, Australia: Sydney University Press, 2008. |
| [19] |
Komeda N, Chaudhary H K, Suzuki G, Mukai Y. Cytological evidence for chromosome elimination in wheat × Imperata cylindrica hybrids. Genes Genet Syst, 2007, 82: 241-248.
doi: 10.1266/ggs.82.241 pmid: 17660694 |
| [20] |
Tayeng T, Chaudhary H K, Kishore N. Enhancing doubled haploid production efficiency in wheat (Triticum aestivum L. em. Thell) by in vivo colchicine manipulations in Imperata cylindrica-mediated chromosome elimination approach. Plant Breed, 2012, 131: 574-578.
doi: 10.1111/pbr.2012.131.issue-5 |
| [21] | Kaila V, Chaudhary H K, Tayeng T, Rather S A. Resolution of genetic mechanism of chromosome elimination in wheat × Imperata cylindrica system of doubled haploidy breeding: the genome responsible. In: Proceedings of National Seminar on Plant Cytogenetics: New Approaches. Patiala, Punjab, India: Department of Botany, Punjabi University, 2012. p 82. |
| [22] | Chaudhary H K. New frontiers in chromosome engineering for enhanced and high precision crop improvement. In: Proceedings of National Seminar on Plant Cytogenetics: New Approaches. Patiala, Punjab, India: Department of Botany, Punjabi University. 2012. pp 35-36. |
| [23] |
Chaudhary H K, Tayeng T, Kaila V, Rather S A. Use of asynchrony in flowering for easy and economical polyhaploid induction in wheat following Imperata cylindrica-mediated chromosome elimination approach. Plant Breed, 2013, 132: 155-158.
doi: 10.1111/pbr.2013.132.issue-2 |
| [24] | Chaudhary H K, Tayeng T, Kaila V, Rather S A. Enhancing the efficiency of wide hybridization mediated chromosome engineering for high precision crop improvement with special reference to wheat × Imperata cylindrica system. Nucleus, 2013, 56: 7-14. |
| [25] |
Rather S A, Chaudhary H K, Kaila V. Proportional contribution and potential of maternal and paternal genotypes for polyhaploid induction in wheat × Imperata cylindrica chromosome elimination approach. Cereal Res Commun, 2014, 42: 19-26.
doi: 10.1556/CRC.2013.0038 |
| [26] |
Mahato A, Chaudhary H K. Relative efficiency of maize and Imperata cylindrica for haploid induction in Triticum durum following chromosome elimination-mediated approach of doubled haploid breeding. Plant Breed, 2015, 134: 379-383.
doi: 10.1111/pbr.2015.134.issue-4 |
| [27] |
Niemirowicz-Szczytt K. Excessive homozygosity in doubled haploids: advantages and disadvantages for plant breeding and fundamental research. Acta Physiol Plant, 1997, 19: 155-167.
doi: 10.1007/s11738-997-0032-7 |
| [28] |
Forster B P, Heberle-Bors E, Kasha K J, Touraev A. The resurgence of haploids in higher plants. Trends Plant Sci, 2007, 12: 368-375.
doi: 10.1016/j.tplants.2007.06.007 pmid: 17629539 |
| [29] | Don Palmer C E, Keller W A. Challenges and Limitations to the Use of Haploidy in crop improvement. In: Don Palmer C E, Keller W A, Kasha K J. Haploids in Crop Improvement II. Berlin, Heidelberg: Springer, 2005. pp 295-303. |
| [30] |
Brownfield L, Kohler C. Unreduced gamete formation in plants: mechanisms and prospects. J Exp Bot, 2011, 62: 1659-1668.
doi: 10.1093/jxb/erq371 pmid: 21109579 |
| [31] |
Xu S J, Dong Y S. Fertility and meiotic mechanisms of hybrids between chromosome autoduplication tetraploid wheats and Aegilops species. Genome, 1992, 35: 379-384.
doi: 10.1139/g92-057 |
| [32] |
Xu S J, Joppa L R. Mechanisms and inheritance of first division restitution in hybrids of wheat, rye, and Aegilops squarrosa. Genome, 1995, 38: 607-615.
pmid: 18470193 |
| [33] |
Matsuoka Y, Nasuda S. Durum wheat as a candidate for the unknown female progenitor of bread wheat: an empirical study with a highly fertile F1 hybrid with Aegilops tauschii Coss. Theor Appl Genet, 2004, 109: 1710-1717.
doi: 10.1007/s00122-004-1806-6 |
| [34] |
Zhang L Q, Chen Q J, Yuan Z W, Xiang Z G, Zheng Y L, Liu D C. Production of aneuhaploid and euhaploid sporocytes by meiotic restitution in fertile hybrids between durum wheat Langdon chromosome substitution lines and Aegilops tauschii. J Genet Genom, 2008, 35: 617-623.
doi: 10.1016/S1673-8527(08)60082-X |
| [35] |
Zhang L Q, Liu D C, Zheng Y L, Yan Z H, Dai S F, Li Y F, Jiang Q, Ye Y Q, Yen Y. Frequent occurrence of unreduced gametes in Triticum turgidum-Aegilops tauschii hybrids. Euphytica, 2010, 172: 285-294.
doi: 10.1007/s10681-009-0081-7 |
| [36] |
Zhang L Q, Yen Y, Zheng Y L, Liu D C. Meiotic restriction in emmer wheat is controlled by one or more nuclear genes that continue to function in derived lines. Sex Plant Reprod, 2007, 20: 159-166.
doi: 10.1007/s00497-007-0052-x |
| [37] |
De Storme N, Copenhaver G P, Geelen D. Production of diploid male gametes in Arabidopsis by cold-induced destabilization of postmeiotic radial microtubule arrays. Plant Physiol, 2012, 160: 1808-1826.
doi: 10.1104/pp.112.208611 pmid: 23096158 |
| [38] |
Zeng D Y, Hao M, Luo J T, Zhang L Q, Yuan Z W, Ning S Z, Zheng Y L, Liu D C. Amphitelic orientation of centromeres at metaphase I is an important feature for univalent-dependent meiotic nonreduction. J Genet, 2014, 93: 531-534.
doi: 10.1007/s12041-014-0393-9 |
| [39] |
Yang Y W, Zhang L Q, Yen Y, Zheng Y L, Liu D C. Cytological evidence on meiotic restitution in pentaploid F1 hybrids between synthetic hexaploid wheat and Aegilops variabilis. Caryologia, 2010, 63: 354-358.
doi: 10.1080/00087114.2010.10589746 |
| [40] | Hao M, Luo J T, Zeng D Y, Zhang L, Ning S Z, Yuan Z W, Yan Z H, Zhang H G, Zheng Y L, Feuillet C, Choulet F, Yen Y, Zhang L Q, Liu D C. QTug.sau-3B is a major quantitative trait locus for wheat hexaploidization. G3: Genes Genom Genet, 2014, 4: 1943-1953. |
| [41] |
寇春兰, 赵来宾, 刘梦, 郝明, 甯顺腙, 袁中伟, 刘登才, 张连全. 小麦未减数配子基因的连锁标记及染色体区段检测. 作物学报, 2016, 42: 984-989.
doi: 10.3724/SP.J.1006.2016.00984 |
|
Kou C L, Zhao L B, Liu M, Hao M, Ning S Z, Yuan Z W, Liu D C, Zhang L Q. Detection of the molecular marker and chromosomal segment linked to unreduced gamete gene in common wheat. Acta Agron Sin, 2016, 42: 984-989. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2016.00984 |
|
| [42] |
Depauw R M, Knox R E, Humphreys D G, Thomas J B, Fox S L, Brown P D, Singh A K, Pozniak C, Randhawa H S, Fowler D B, Graf R J, Hucl P. New breeding tools impact Canadian commercial farmer fields. Czech J Genet Plant Breed, 2011, 47: S28-S34.
doi: 10.17221/3250-CJGPB |
| [43] | Pauk J, Mihály R, Puolimatka M. Protocol for wheat (Triticum aestivum L.) anther culture. In: Maluszynski M, Kasha K J, Forster B P, Szarejko I, eds. Doubled Haploid Production in Crop Plants. Dordrecht: Springer Netherlands, 2003. pp 59-64. |
| [44] | Patial M, Pal D, Thakur A, Bana R S, Patial S. Doubled haploidy techniques in wheat (Triticum aestivum L.): an overview. Natl Acad Sci India Proc, Section B: Biol Sci, 2019, 89: 27-41. |
| [1] | 寇春兰,赵来宾,刘梦,郝明,甯顺腙,袁中伟,刘登才,张连全*. 小麦未减数配子基因的连锁标记及染色体区段检测[J]. 作物学报, 2016, 42(07): 984-989. |
| [2] | 许树军;董玉琛. 波斯小麦×节节麦杂种F_1直接形成双二倍体的细胞遗传学研究[J]. 作物学报, 1989, 15(03): 251-259. |
|