强再生力水稻品种碳氮营养与激素生理特征研究
黄素华, 林席跃, 雷正平, 丁在松, 赵明
作物学报
2021, 47 ( 11):
2278-2289.
DOI: 10.3724/SP.J.1006.2021.02070
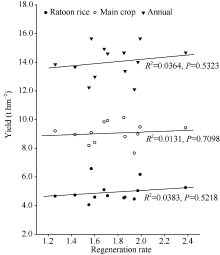
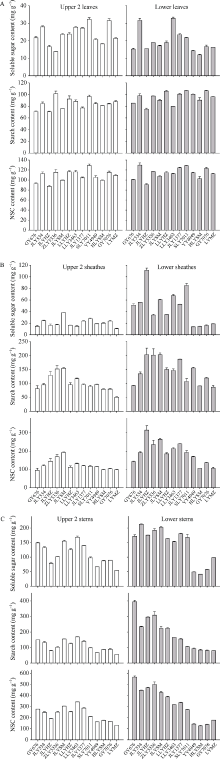
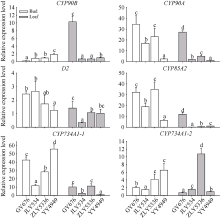
明确强再生力品种腋芽萌发的生理基础与激素调控特点对于再生稻品种筛选和栽培技术调控具有重要意义。本研究利用在江西崇义县建立的再生稻品种筛选平台, 分析了2019年筛选的13个品种头季收获时不同部位的非结构性碳水化合物(non-structural carbohydrate, NSC)及全氮含量, 并对促进和抑制腋芽萌发的主要激素油菜素内酯和独脚金内酯的合成和信号转导关键基因的表达进行了研究。结果表明, 13个品种的再生力存在较大的差异, 变化范围为1.26~2.38; 不同品种之间, 不同节位之间的叶片、叶鞘和茎秆的可溶性糖、淀粉和非结构性碳水化合物含量均存在极显著的差异(P值均小于0.001); 而全氮含量除了上下节位茎秆的差异不显著外, 其余的也均存在极显著的差异; 与再生力的相关性分析表明仅有下部节位茎秆的可溶性糖、淀粉和NSC含量与再生力相关性达到显著或极显著水平(R2分别为0.4442*、0.9000**和0.8303**), 而其他均无显著相关性。强再生力品种谷优676中BR合成和信号途径中促进分蘖的基因CYP90A、CYP852A、D2、BRI、BSK和CYCD3表达水平增高, 而抑制分蘖的基因CYP734A1、BZR和BKI表达水平较低。可见, 可以利用下部节位茎秆的淀粉含量作为强再生力品种的筛选指标, 同时以BR途径相关基因表达水平作为辅助指标。
器官
Organ | 差异源
Source of variation | 自由度
df | 可溶性糖SS | 淀粉Starch | 非结构性碳水化合物NSC | | F | P | F | P | F | P | | 叶片Leaf | 节位 Node site | 1 | 2113.78 | 8.66E-44 | 1727.37 | 1.44E-41 | 594.95 | 3.92E-30 | | 品种 Cultivar | 12 | 854.64 | 3.12E-55 | 387.53 | 2.20E-46 | 497.91 | 3.51E-49 | | 叶鞘Sheath | 节位 Node site | 1 | 12,932.43 | 5.13E-64 | 832.86 | 1.13E-33 | 2473.73 | 1.59E-45 | | 品种 Cultivar | 12 | 1391.03 | 1.05E-60 | 173.85 | 1.63E-37 | 308.29 | 7.72E-44 | | 茎秆Stem | 节位 Node site | 1 | 3384.81 | 5.26E-49 | 1396.50 | 3.03E-39 | 3923.92 | 1.19E-50 | | 品种 Cultivar | 12 | 2564.40 | 1.35E-67 | 1476.25 | 2.24E-61 | 3189.50 | 4.71E-70 |
View table in article
表4
不同水稻品种不同节位间可溶性糖、淀粉和NSC含量方差分析
正文中引用本图/表的段落
早期的研究表明, 强再生力品种头季稻的农艺性状主要表现为分蘖力强、有效穗多、穗粒数偏少、齐穗期叶粒比大的特征[4,5]。任天举等[6]对229个水稻杂交组合再生力与农艺性状的通径分析指出成熟期单茎的茎鞘干重对再生力的直接效应最大或接近最大, 可将茎鞘干重作为衡量品种再生力的重要指标[7,8]。进一步的研究发现头季稻收获后稻桩的干物质量与再生力和再生季产量具有显著的相关性[9]。通过去除部分叶片和/或小穗对头季稻进行源库调节的研究也表明其对再生季再生力的影响也与稻桩的干物质及非结构性碳水化合物(non-structural carbohydrates, NSC)含量的改变有关[10]。栽培技术调控的研究表明适时施用促芽肥是提高再生稻再生力的有效技术措施[11,12], 头季收获后使用促苗肥并进行土壤干湿交替的水分管理可以通过促进再生季根系生长和腋芽萌发提高再生力[13]。徐富贤等[1]认为其机制在于齐穗后施用氮肥延缓了叶片衰老, 提高了叶片光合物质生产, 从而增加了母茎茎鞘的干物质积累。可见, 茎鞘干物质很可能是其中储藏的养分物质对再生芽的发育起着重要作用。然而, 对再生稻头季收获时的碳、氮养分及其在不同器官(叶、茎和鞘), 不同节位的分布特征尚缺乏深入分析。另外, 近年来研究表明油菜素内酯(brassinosteroids, BRs)和独脚金内酯(strigolactones, SLs)两种激素信号分别在促进和抑制水稻分蘖发生中起着重要作用[14,15], 而在强再生力水稻品种再生芽的萌发过程中, 这2种激素的合成和信号转导途径是否也参与调控还未见报道。
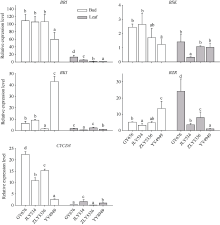
图2显示了13个水稻品种头季收获前植株上、下不同节位叶片、叶鞘和茎秆的可溶性糖(soluble sugar, SS), 淀粉和NSC含量。叶片中SS的变化范围分别为13.9~32.3 mg g-1和12.0~32.9 mg g-1, 淀粉含量的变化范围分别为70.9~101.5 mg g-1和75.3~106.8 mg g-1, NSC含量变化范围为87.6~129.1 mg g-1和90.8~129.9 mg g-1; 叶鞘中SS含量变化范围分别为10.6~38.5 mg g-1和13.8~111.4 mg g-1, 淀粉含量的变化范围分别为50.7~154.7 mg g-1和87.1~203.8 mg g-1, NSC含量变化范围为95.8~193.2 mg g-1和105.9~315.2 mg g-1; 茎秆中SS含量变化范围分别为54.8~168.6 mg g-1和41.4~213.0 mg g-1, 淀粉含量的变化范围分别为75.9~173.5 mg g-1和79.3~394.2 mg g-1, NSC含量变化范围为109.5~304.7 mg g-1和125.3~565.9 mg g-1。方差分析表明品种间以及上下节位间的SS、淀粉和NSC含量均存在极显著的差异(表4)。各个组分在上、下部节位叶片之间的差异明显小于叶鞘和茎秆。
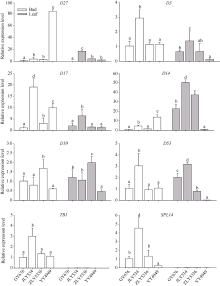
如图7所示, 水稻中参与SL合成的D27、D17和D10中, 合成途径上游的基因D27在甬优4949腋芽中具有极高的表达量, 是其他3个高再生率品种中表达量的30倍以上, 而合成途径中间和下游的D17和D10的表达量与再生率没有明显的相关特征。SL信号转导途径中的D3、D14、D53、SPL14和TB1基因, 除D14在4个品种中的表达特征比较特异外, 其他4个基因在晶两优534腋芽中的表达明显高于另外3个品种。而在叶片中, TB1和SPL14表达量极低, 几乎检测不到; D3、D14和D53的表达在再生率高的3个品种中高于甬优4949。
本文的其它图/表
|