欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (10): 2425-2434.doi: 10.3724/SP.J.1006.2024.43012
• 作物遗传育种·种质资源·分子遗传学 • 下一篇
黄灵芝1,2( ), 符晓2, 祁显涛2, 刘昌林2, 谢传晓2, 吴鹏昊1, 任姣姣1,*(), 朱金洁2,*()
), 符晓2, 祁显涛2, 刘昌林2, 谢传晓2, 吴鹏昊1, 任姣姣1,*(), 朱金洁2,*()
HUANG Ling-Zhi1,2(), FU Xiao2, QI Xian-Tao2, LIU Chang-Lin2, XIE Chuan-Xiao2, WU Peng-Hao1, REN Jiao-Jiao1,*(), ZHU Jin-Jie2,*()
摘要:
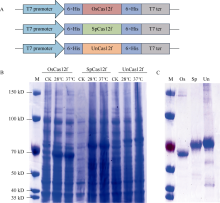
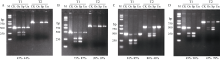
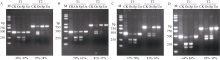
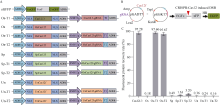
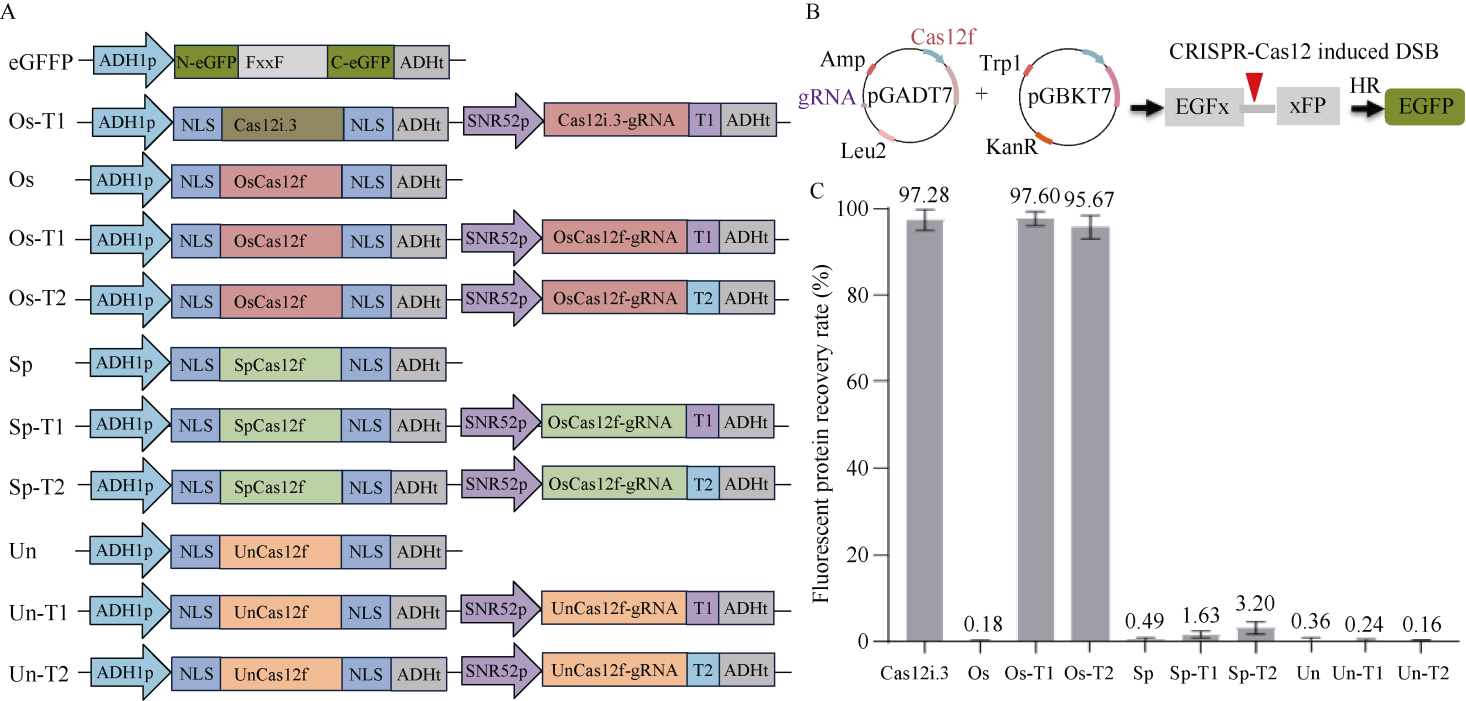
来自Type V-F家族的CRISPR/Cas12f蛋白报道仅为Cas9蛋白分子的1/4到1/3大小, 在病毒载体的递送上具有重要优势。然而, CRISPR/Cas12f系统介导植物基因编辑的报道较少, 编辑活性相对较低, 限制了该系统在植物上的进一步应用。本研究分别在体外酶切、酵母以及玉米原生质体瞬时表达3个体系中比较了OsCas12f、SpCas12f及UnCas12f的编辑活性。 结果表明, 基于Cas12f/sgRNA的体外酶切, OsCas12f与SpCas12f蛋白的编辑活性相当, 未检测到UnCas12f对底物酶切活性; 在酵母突变eGFP的恢复表达试验中, OsCas12f在2个测试位点对eGFP蛋白的表达恢复效率达到95%以上, 效率与Cas12i.3相当; SpCas12f介导的2个位点eGFP蛋白表达恢复效率分别是1.63%与3.20%, 效果次之; UnCas12f蛋白几乎无编辑活性; 玉米原生质体瞬时表达比较OsCas12f和SpCas12f介导的玉米内源位点的编辑效率, 发现OsCas12f对2个位点的编辑效率分别为2.72%和1.97%, 而SpCas12f仅能介导其中1个位点的定点编辑, 编辑效率为1.09%。Cas12f蛋白在靶位点处引入的突变类型以碱基的缺失为主, 缺失碱基长度在-9~ -17 bp之间。综上, OsCas12f可以作为植物微型基因编辑器及衍生技术开发的底盘工具酶。
| [1] |
Chen K, Wang Y, Zhang R, Zhang H, Gao C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu Rev Plant Biol, 2019, 70: 667-697.
doi: 10.1146/annurev-arplant-050718-100049 pmid: 30835493 |
| [2] |
Geurts A M, Cost G J, Freyvert Y, Zeitler B, Miller J C, Choi VM, Jenkins S S, Wood A, Cui X, Meng X, Vincent A, Lam S, Michalkiewicz M, Schilling R, Foeckler J, Kalloway S, Weiler H, Ménoret S, Anegon I, Davis G D, Zhang L, Rebar E J, Gregory P D, Urnov F D, Jacob H J, Buelow R. Knockout rats via embryo microinjection of zinc-finger nucleases. Science, 2009, 325: 433.
doi: 10.1126/science.1172447 pmid: 19628861 |
| [3] | An Y, Geng Y, Yao J, Fu C, Lu M, Wang C, Du J. Efficient genome editing in populus using CRISPR/Cas12a. Front Plant Sci, 2020, 11: 593938. |
| [4] |
Makarova K S, Wolf Y I, Alkhnbashi O S, Costa F, Shah S A, Saunders S J, Barrangou R, Brouns S J, Charpentier E, Haft D H, Horvath P, Moineau S, Mojica F J, Terns R M, Terns M P, White M F, Yakunin A F, Garrett R A, van der Oost J, Backofen R, Koonin E V. An updated evolutionary classification of CRISPR-Cas systems. Nat Rev Microbiol, 2015, 13: 722-736.
doi: 10.1038/nrmicro3569 pmid: 26411297 |
| [5] |
Makarova K S, Wolf Y I, Iranzo J, Shmakov S A, Alkhnbashi O S, Brouns S J J, Charpentier E, Cheng D, Haft D H, Horvath P, Moineau S, Mojica F J M, Scott D, Shah S A, Siksnys V, Terns M P, Venclovas Č, White M F, Yakunin A F, Yan W, Zhang F, Garrett R A, Backofen R, van der Oost J, Barrangou R, Koonin E V. Evolutionary classification of CRISPR-Cas systems: a burst of class 2 and derived variants. Nat Rev Microbiol, 2020, 18: 67-83.
doi: 10.1038/s41579-019-0299-x pmid: 31857715 |
| [6] |
Nishimasu H, Cong L, Yan W X, Ran F A, Zetsche B, Li Y, Kurabayashi A, Ishitani R, Zhang F, Nureki O. Crystal Structure of Staphylococcus aureus Cas9. Cell, 2015, 162: 1113-1126.
doi: 10.1016/j.cell.2015.08.007 pmid: 26317473 |
| [7] | Kim E, Koo T, Park S W, Kim D, Kim K, Cho H Y, Song D W, Lee K J, Jung M H, Kim S, Kim J H, Kim J H, Kim J S. In vivo genome editing with a small Cas 9 orthologue derived from Campylobacter jejuni. Nat Commun, 2017, 8: 14500. |
| [8] |
Zetsche B, Gootenberg J S, Abudayyeh O O, Slaymaker I M, Makarova K S, Essletzbichler P, Volz S E, Joung J, van der Oost J, Regev A, Koonin E V, Zhang F. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell, 2015, 163: 759-771.
doi: 10.1016/j.cell.2015.09.038 pmid: 26422227 |
| [9] |
Teng F, Cui T, Gao Q, Guo L, Zhou Q, Li W. Artificial sgRNAs engineered for genome editing with new Cas12b orthologs. Cell Discov, 2019, 5: 23.
doi: 10.1038/s41421-019-0091-0 pmid: 31016029 |
| [10] |
McGaw C, Garrity A J, Munoz G Z, Haswell J R, Sengupta S, Keston-Smith E, Hunnewell P, Ornstein A, Bose M, Wessells Q, Jakimo N, Yan P, Zhang H, Alfonse L E, Ziblat R, Carte J M, Lu W C, Cerchione D, Hilbert B, Sothiselvam S, Yan W X, Cheng D R, Scott D A, DiTommaso T, Chong S. Engineered Cas12i2 is a versatile high-efficiency platform for therapeutic genome editing. Nat Commun, 2022, 13: 2833.
doi: 10.1038/s41467-022-30465-7 pmid: 35595757 |
| [11] | Wang Y, Qi T, Liu J, Yang Y, Wang Z, Wang Y, Wang T, Li M, Li M, Lu D, Chang A C Y, Yang L, Gao S, Wang Y, Lan F. A highly specific CRISPR-Cas12j nuclease enables allele-specific genome editing. Sci Adv, 2023, 9: eabo6405. |
| [12] | Hirano S, Kappel K, Altae-Tran H, Faure G, Wilkinson M E, Kannan S, Demircioglu F E, Yan R, Shiozaki M, Yu Z, Makarova K S, Koonin E V, Macrae R K, Zhang F. Structure of the OMEGA nickase IsrB in complex with ωRNA and target DNA. Nature, 2022, 610: 575-581. |
| [13] |
Altae-Tran H, Kannan S, Demircioglu F E, Oshiro R, Nety S P, McKay L J, Dlakić M, Inskeep W P, Makarova K S, Macrae R K, Koonin E V, Zhang F. The widespread IS200/IS605 transposon family encodes diverse programmable RNA-guided endonucleases. Science, 2021, 374: 57-65.
doi: 10.1126/science.abj6856 pmid: 34591643 |
| [14] | Kim D Y, Lee J M, Moon S B, Chin H J, Park S, Lim Y, Kim D, Koo T, Ko J H, Kim Y S. Efficient CRISPR editing with a hypercompact Cas12f1 and engineered guide RNAs delivered by adeno-associated virus. Nat Biotechnol, 2022, 40: 94-102. |
| [15] |
Xin C, Yin J, Yuan S, Ou L, Liu M, Zhang W, Hu J. Comprehensive assessment of miniature CRISPR-Cas12f nucleases for gene disruption. Nat Commun, 2022, 13: 5623.
doi: 10.1038/s41467-022-33346-1 pmid: 36153319 |
| [16] | Wang Y, Wang Y, Pan D, Yu H, Zhang Y, Chen W, Li F, Wu Z, Ji Q. Guide RNA engineering enables efficient CRISPR editing with a miniature Syntrophomonas palmitatica Cas12f1 nuclease. Cell Rep, 2022, 40: 111418. |
| [17] |
Kong X, Zhang H, Li G, Wang Z, Kong X, Wang L, Xue M, Zhang W, Wang Y, Lin J, Zhou J, Shen X, Wei Y, Zhong N, Bai W, Yuan Y, Shi L, Zhou Y, Yang H. Engineered CRISPR-OsCas12f1 and RhCas12f1 with robust activities and expanded target range for genome editing. Nat Commun, 2023, 14: 2046.
doi: 10.1038/s41467-023-37829-7 pmid: 37041195 |
| [18] | Sukegawa S, Nureki O, Toki S, Saika H. Genome editing in rice mediated by miniature size Cas nuclease SpCas12f. Front Genome Ed, 2023, 5: 1138843. |
| [19] | Jiang Y Y, Chai Y P, Lu M H, Han X L, Lin Q, Zhang Y, Zhang Q, Zhou Y, Wang X C, Gao C, Chen Q J. Prime editing efficiently generates W542L and S621I double mutations in two ALS genes in maize. Genome Biol, 2020, 21: 257. |
| [20] | Bähler J, Wise J A. Preparation of total RNA from fission yeast. Cold Spring Harb Protoc, 2017, 2017: pdb. prot091629. |
| [21] | Cao J, Yao D, Lin F, Jiang M Y. PEG-mediated transient gene expression and silencing system in maize mesophyll protoplasts: a valuable tool for signal transduction study in maize. Acta Physiol Plant, 2014, 36: 1271-1281. |
| [22] |
Karvelis T, Bigelyte G, Young J K, Hou Z, Zedaveinyte R, Budre K, Paulraj S, Djukanovic V, Gasior S, Silanskas A, Venclovas Č, Siksnys V. PAM recognition by miniature CRISPR-Cas12f nucleases triggers programmable double-stranded DNA target cleavage. Nucleic Acids Res, 2020, 48: 5016-5023.
doi: 10.1093/nar/gkaa208 pmid: 32246713 |
| [23] | Xiao R, Li Z, Wang S, Han R, Chang L. Structural basis for substrate recognition and cleavage by the dimerization-dependent CRISPR-Cas12f nuclease. Nucleic Acids Res, 2021, 49: 4120-4128. |
| [24] |
Bigelyte G, Young J K, Karvelis T. Miniature type V-F CRISPR- Cas nucleases enable targeted DNA modification in cells. Nat Commun, 2021, 12: 6191.
doi: 10.1038/s41467-021-26469-4 pmid: 34702830 |
| [25] |
Hill Z B, Martinko A J, Nguyen D P, Wells J A. Human antibody-based chemically induced dimerizers for cell therapeutic applications. Nat Chem Biol, 2018, 14: 112-117.
doi: 10.1038/nchembio.2529 pmid: 29200207 |
| [1] | 莫广玲, 余陈静, 梁艳兰, 周定港, 罗俊, 王莫, 阙友雄, 黄宁, 凌辉. 甘蔗ScbHLH13基因的RT-PCR克隆与功能分析[J]. 作物学报, 2023, 49(9): 2485-2497. |
| [2] | 杨梦婷, 张春, 王作平, 邹华文, 吴忠义. 玉米ZmbHLH161基因的克隆及功能研究[J]. 作物学报, 2020, 46(12): 2008-2016. |
| [3] | 殷龙飞,王朝阳,吴忠义,张中保,于荣. 玉米ZmGRAS31基因的克隆及功能研究[J]. 作物学报, 2019, 45(7): 1029-1037. |
| [4] | 李继洋,胡燕,姚瑞,代培红,刘晓东. 基于优化sgRNA系统提高海岛棉CRISPR/Cas9基因组编辑功效的研究[J]. 作物学报, 2019, 45(10): 1522-1534. |
| [5] | 彭章, 童华荣, 梁国鲁, 石艺琦, 袁连玉. 茶树叶片和胚根原生质体的分离及PEG诱导融合[J]. 作物学报, 2018, 44(03): 463-470. |
| [6] | 李继洋, 雷建峰, 代培红, 姚瑞, 曲延英, 陈全家, 李月, 刘晓东. 基于棉花U6启动子的海岛棉CRISPR/Cas9基因组编辑体系的建立[J]. 作物学报, 2018, 44(02): 227-235. |
| [7] | 郭艳萍,任成杰,李志伟,王文斌,张仁和,路海东,刘建超,张兴华,薛吉全,郭东伟. 玉米胚乳细胞原生质体的分离与流式纯化[J]. 作物学报, 2014, 40(03): 424-430. |
| [8] | 李妮娜,丁林云,张志远,郭旺珍. 棉花叶肉原生质体分离及目标基因瞬时表达体系的建立[J]. 作物学报, 2014, 40(02): 231-239. |
| [9] | 胡琼;李云昌. 体细胞杂交在油菜细胞质雄性不育创建和改良中的应用[J]. 作物学报, 2006, 32(01): 138-143. |
| [10] | 李韬;戴朝. 提高马铃薯原生质体细胞分裂频率的研究[J]. 作物学报, 2000, 26(06): 953-958. |
| [11] | 杨跃生;简玉瑜;陈远玲. 水稻原生质体愈伤组织再生植株培养程序的比较[J]. 作物学报, 2000, 26(04): 490-495. |
| [12] | 肖文言;王连铮. 大豆幼荚子叶原生质体培养及植株再生[J]. 作物学报, 1994, 20(06): 665-669. |
| [13] | 鲁雪华;刘中柱;林永烈;林大添;丁舒敏. 旱稻原生质体培养和植株再生的研究[J]. 作物学报, 1994, 20(04): 419-425. |
| [14] | 刘宝;吴琴生;刘大钧. 大麦原生质体培养再生胚性愈伤组织和白化苗[J]. 作物学报, 1993, 19(06): 520-522. |
| [15] | 吕慧能;盖钧镒;马育华;卫志明;许智宏. 不同激素条件下大豆原生质体培养和植株再生[J]. 作物学报, 1993, 19(04): 328-333. |
|