欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (6): 1100-1108.doi: 10.3724/SP.J.1006.2021.04138
黄冰艳1( ), 孙子淇1, 刘华1, 房元瑾1, 石磊1, 苗利娟1, 张毛宁1, 张忠信1, 徐静1, 张梦圆2, 董文召1, 张新友1,*()
), 孙子淇1, 刘华1, 房元瑾1, 石磊1, 苗利娟1, 张毛宁1, 张忠信1, 徐静1, 张梦圆2, 董文召1, 张新友1,*()
HUANG Bing-Yan1(), SUN Zi-Qi1, LIU Hua1, FANG Yuan-Jin1, SHI Lei1, MIAO Li-Juan1, ZHANG Mao-Ning1, ZHANG Zhong-Xin1, XU Jing1, ZHANG Meng-Yuan2, DONG Wen-Zhao1, ZHANG Xin-You1,*()
摘要:
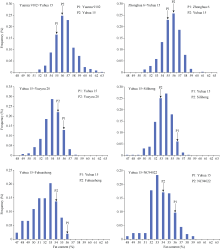
巢式群体可以利用多个亲本解析复杂性状的遗传机制。本研究利用1个共同亲本与6个基础亲本所配置巢式组合F2:3家系的种子脂肪含量数据, 分析了花生脂肪含量的遗传模型, 旨在探明不同的基础亲本组合中脂肪含量性状的遗传差异, 为制定脂肪含量遗传改良的亲本选配和后代选择策略提供依据。6个组合的共同亲本为高脂肪含量的普通型大果品种豫花15号, 其他6个基础亲本为不同脂肪含量和不同植物学类型的品种。结果表明, 在不同杂交组合中脂肪含量的遗传模式有所不同, 6个组合分别符合无主基因模型、1对主基因加性显性模型和2对主基因等显性模型3种遗传模式。各种遗传效应的估计值也各不相同, 主基因遗传力从32%到80%, 说明不同杂交组合中, 控制脂肪含量的基因位点差异及其重组和分离方式不同。高脂肪含量双亲杂交后代的高脂肪含量个体较多, 但主基因遗传力较低, 不宜在早代实施表型选择; 双亲脂肪含量差异较大的后代脂肪含量变异幅度更大, 能够选择到不同脂肪含量的类型。本研究也表明, 巢式组合具有较丰富的脂肪含量变异类型, 揭示出脂肪含量性状遗传的复杂性和多基因调控的特点, 为较全面地了解脂肪含量的遗传提供了基础。该巢式群体也将有助于进一步开展脂肪含量的QTL定位研究。
| [1] | FAOSTAT, 2020. FAO. http://www.fao.org/faostat/en/#home. |
| [2] | 廖伯寿, 雷永, 王圣玉, 李栋, 黄家权, 姜慧芳, 任小平. 花生重组近交系群体的遗传变异与高油种质的创新. 作物学报, 2008,34:999-1004. |
| Liao B S, Lei Y, Wang S Y, Li D, Huang J Q, Jiang H F, Ren X P. Genetic diversity of peanut RILs and enhancement for high oil genotypes. Acta Agron Sin, 2008,34:999-1004 (in Chinese with English abstract). | |
| [3] | 禹山林, 杨庆利, 潘丽娟, 薄文娜. 花生种子含油量的遗传分析. 植物遗传资源学报, 2009,10:453-456. |
| Yu S L, Yang Q L, Pan L J, Bo W N. Genetic analysis for oil content of peanut seeds. J Plant Genet Resour, 2009,10:453-456 (in Chinese with English abstract). | |
| [4] | 陈四龙, 李玉荣, 程增书, 廖伯寿, 雷永, 刘吉生. 花生含油量杂种优势表现及主基因+多基因遗传效应分析. 中国农业科学, 2009,42:3048-3057. |
| Chen S L, Li Y R, Cheng Z S, Liao B S, Lei Y, Liu J S. Heterosis and genetic analysis of oil content in peanut using mixed model of major gene and polygene. Sci Agric Sin, 2009,42:3048-3057 (in Chinese with English abstract). | |
| [5] | Zhang X, Zhu S, Han S, Xu J, Liu H, Tang F, Dong W, Zang X, Zhang Z X. Inheritance of fat and fatty acid compositions in peanut (Arachis hypogaea L.). Agric Sci Technol, 2011,12:943-946. |
| [6] | 刘华, 张新友, 崔党群, 汤丰收, 董文召, 韩锁义, 徐静, 臧秀旺, 张忠信. 花生蛋白质和脂肪含量的主基因+多基因遗传分析. 江苏农业科学, 2011,39(2):127-130. |
| Liu H, Zhang X Y, Cui D X, Tang F S, Dong W Z, Han S Y, Xu J, Zang X W, Zhang Z X. Genetic analysis of protein and fat content using major gene plus polygene methods in peanut (Arachis hypogaea L.). Jiangsu Agric Sci, 2011,39(2):127-130 (in Chinese with English abstract). | |
| [7] | 刘华, 秦利, 张新友, 韩锁义, 杜培, 张忠信, 孙子淇, 齐飞艳, 董文召, 黄冰艳. 基于花生种间杂交遗传群体的脂肪及脂肪酸含量的遗传模型分析. 中国油料作物学报, 2016,38:172-178. |
| Liu H, Qin L, Zhang X Y, Han S Y, Du P, Zhang Z X, Sun Z Q, Qi F Y, Dong W Z, Huang B Y. Genetic models of peanut fat and fatty acid compositions based on interspecific RIL population. Chin J Oil Crop Sci, 2016,38:172-178 (in Chinese with English abstract). | |
| [8] | 王建康, 盖钧镒. 利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应. 遗传学报, 1997,24:432-440. |
| Wang J K, Gai J Y. Identification of major gene and polygene mixed inheritance model and estimation of genetic parameters of a quantitative trait from F2 progeny. Act Genet Sin, 1997,24:432-440 (in Chinese with English abstract). | |
| [9] | Meng L, Li H, Zhang L, Wang J. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in bi-parental populations. Crop J, 2015,3:269-283. |
| [10] | 甘信民, 曹玉良, 魏家祥, 崔务峰, 顾淑媛, 刘法生. 花生数量性状配合力的研究. 中国油料作物学报, 1981, (3):33-45. |
| Gan S M, Cao Y L, Wei J X, Cui W F, Gu S Y, Liu F S. Combining ability analysis of quatitative traits in peanut. Chin J Oil Crop Sci, 1981, ( 3):33-45 (in Chinese with English abstract). | |
| [11] | Isleib T G, Pattee H E, Giesbrecht F G. Oil, sugar, and starch characteristics in peanut breeding lines selected for low and high oil content and their combining ability. J Agric Food Chem, , 200452:3165-3168. |
| [12] | 徐宜民, 甘信民, 曹玉良, 顾淑媛, 刘法生. 花生主要营养品质性状和农艺性状配合力的研究. 中国农业科学, 1995,28(2):15-23. |
| Xu Y M, Gan X M, Cao Y L, Gu S Y, Liu F S. studies on combining ability of major nutritional quality characters and agronomic characters in peanut. Sci Agric Sin, 1995,28(2):15-23 (in Chinese with English abstract) . | |
| [13] | 梁炫强, 郑广柔, 向荣英, 黎秀英. 珍珠豆型花生产量和含油率性状配合力分析. 花生科技, 1991, ( 3):11-14. |
| Liang X Q, Zheng G R, Xiang R Y, Li X Y. Combining ability analysis of yield and oil content of Spanish type peanut. Sci Technol Peanut, 1991, ( 3):11-14 (in Chinese with English abstract). | |
| [14] | 梁炫强, 郑广柔, 向荣英, 黎秀英. 珍珠豆型花生产量和含油率遗传特性的双列分析. 中国油料, 1992, ( 1):19-23. |
| Liang X Q, Zheng G R, Xiang R Y, Li X Y. A diallel analysis of yield and oil content of spanish type peanut. Chin Oil Crop, 1992, ( 1):19-23 (in Chinese with English abstract). | |
| [15] | 付三雄, 戚存扣. 甘蓝型油菜含油量的主基因+多基因遗传分析. 江苏农业学报, 2009,25:731-736. |
| Fu S X, Qi C K. Major gene plus polygene inheritance of oil content in Brassica napus L. Jiangsu J Agric Sci, 2009,25:731-736 (in Chinese with English abstract). | |
| [16] | Zhang S F, Ma C Z, Zhu J C, Wang J P, Wen Y C, Fu T D. Genetic analysis of oil content in Brassica napus L. using mixed model of major gene and polygene. Acta Genet Sin, 2006,2:171-180. |
| [17] | 金梦阳, 李加纳, 付福友, 张正圣, 张学昆, 刘列钊. 甘蓝型油菜含油量及皮壳率的QTL分析. 中国农业科学, 2007,40:677-684. |
| Jin M Y, Li J N, Fu F Y, Zhang Z S, Zhang X K, Liu L Z. QTL analysis of oil and hull content in Brassica napus L. Sci Agric Sin, 2007,40:677-684 (in Chinese with English abstract). | |
| [18] | 张洁夫, 戚存扣, 浦惠明, 陈松, 陈锋, 高建芹, 陈新军, 顾慧, 傅寿仲. 甘蓝型油菜含油量的遗传与QTL定位. 作物学报, 2007,33:1495-1501. |
| Zhang J F, Qi C K, Pu H M, Chen S, Chen F, Gao J Q, Chen X J, Gu H, Fu S Z. Inheritance and QTL identification of oil content in rapeseed (Brassica napus L.). Acta Agron Sin, 2007,33:1495-1501 (in Chinese with English abstract). | |
| [19] | 张新友, 韩锁义, 徐静, 严玫, 刘华, 汤丰收, 董文召, 黄冰艳. 花生主要品质性状的QTLs定位分析. 中国油料作物学报, 2012,34:311-315. |
| Zhang X Y, Han S Y, Xu J, Yan M, Liu H, Tang F S, Dong W Z, Huang B Y. Identification of QTLs for important quality traits in cultivated peanut (Arachis hypogaea L.). Chin J Oil Crop Sci, 2012,34:311-315 (in Chinese with English abstract). | |
| [20] | 黄莉, 赵新燕, 张文华, 樊志明, 任小平, 廖伯寿, 姜慧芳, 陈玉宁. 利用RIL群体和自然群体检测与花生含油量相关的SSR标记. 作物学报, 2011,37:1967-1974. |
| Huang L, Zhao X Y, Zhang W H, Fan Z M, Ren X P, Liao B S, Jiang H F, Chen Y N. Identification of SSR markers linked to oil content in peanut (Arachis hypogaea L.) through RIL population and natural population. Acta Agron Sin, 2011,37:1967-1974 (in Chinese with English abstract). | |
| [21] | Pandey M K, Wang M L, Qiao L X, Feng S P, Khera P, Wang H, Tonnis B, Barkley N A, Wang J P, Holbrook C C, Culbreath A K, Varshney R K, Guo B Z. Identification of QTLs associated with oil content and mapping FAD2 genes and their relative contribution to oil quality in peanut (Arachis hypogaea L.). BMC Genet, 2014,15:133. |
| [22] | 魏大勇, 崔艺馨, 梅家琴, 汤青林, 李加纳, 钱伟. 油菜种子含油量GWAS分析及位点整合系统构建. 作物学报, 2018,44:1311-1319. |
| Wei D Y, Cui Y X, Mei J Q, Tang Q L, Li J N, Qian W. Genome-wide association study on seed oil content in rapeseed and construction of integration system for oil content loci. Acta Agron Sin, 2018,44:1311-1319 (in Chinese with English abstract). | |
| [23] |
Sun F M, Liu J, Hua W, Sun X C, Wang X F, Wang H Z. Identification of stable QTLs for seed oil content by combined linkage and association mapping in Brassica napus. Plant Sci, 2016,252:388-399.
pmid: 27717475 |
| [24] | Yu J, Holland J B, McMullen M D, Buckler E S. Genetic design and statistical power of nested association mapping in maize. Genetics, 2008,178:539-551. |
| [25] | Li H, Bradbury P, Ersoz E, Buckler E S, Wang J. Joint QTL linkage mapping for multiple-cross mating design sharing one common parent. PLoS One, 2011,6:e17573. |
| [26] | Xavier A, Xu S, Muir W M, Rainey K M. NAM: association studies in multiple populations. Bioinformatics, 2015,31:3862-3864. |
| [27] | Bouchet S, Olatoye M O, Marla SR, Perumal R, Tesso T, Yu J M, Tuinstra M, Morris G P. Increased power to dissect adaptive traits in global sorghum diversity using a nested association mapping population. Genetics, 2017,206:573-585. |
| [28] | Schnaithmann F, Kopahnke D, Pillen K. A first step toward the development of a barley NAM population and its utilization to detect QTLs conferring leaf rust seedling resistance. Theor Appl Genet, 2014,127:1513-1525. |
| [29] | Xavier A, Jarquin D, Howard R, Ramasubramanian V, Specht J E, Graef G L, Beavis W D, Diers B W, Song Q, Cregan P B, Nelson R, Mian R, Shannon J G, McHale L, Wang D, Schapaugh W, Lorenz A J, Xu S, Muir W M., Rainey K M. Genome-wide analysis of grain yield stability and environmental interactions in a multiparental soybean population. G3: Genes Genom Genet, 2018,8:519-529. |
| [30] | Khan M A, Tong F, Wang W, He J, Zhao T, Gai J. Analysis of QTL-allele system conferring drought tolerance at seedling stage in a nested association mapping population of soybean [ Glycine max (L.) Merr.] using a novel GWAS procedure. Planta, 2018,248:947-962. |
| [31] | Chen Q, Yang C J, York A M, Xue W, Daskalska L L, DeValk C A, Krueger K W, Lawton S B, Spiegelberg B G, Schnell J M, Neumeyer M A, Perry J S, Peterson A C, Kim B, Bergstrom L, Yang L, Barber I C, Tian F, Doebley J F. TeoNAM: a nested association mapping population for domestication and agronomic trait analysis in maize. Genetics, 2019,213:1065-1078. |
| [32] | Beche E, Gillman J D, Song Q, Nelson R, Beissinger T, Decker J, Shannon G, Scaboo A M. Nested association mapping of important agronomic traits in three interspecific soybean populations. Theor Appl Genet, 2020,133:1039-1054. |
| [33] |
Garin V, Wimmer V, Mezmouk S, Malosetti M, Eeuwijk F. How do the type of QTL effect and the form of the residual term influence QTL detection in multi-parent populations? A case study in the maize EU-NAM population. Theor Appl Genet, 2017,130:1753-1764.
pmid: 28547012 |
| [34] | Monir M M, Zhu J. Dominance and epistasis interactions revealed as important variants for leaf traits of maize NAM population. Front Plant Sci, 2018,9:627. |
| [35] |
Hu J, Guo C, Wang B, Ye J, Liu M, Wu Z, Xiao Y, Zhang Q, Li H, King G J, Liu K. Genetic properties of a nested association mapping population constructed with semi-winter and spring oilseed rapes. Front Plant Sci, 2018,9:1740.
pmid: 30534135 |
| [36] | Jordan K W, Wang S, He F, Chao S, Lun Y, Paux E, Sourdille P, Sherman J, Akhunova A, Blake N K, Pumphrey M O, Glover K, Dubcovsky J, Talbert L, Akhunov E D. The genetic architecture of genome-wide recombination rate variation in allopolyploid wheat revealed by nested association mapping. Plant J, 2018,95:1039-1054. |
| [37] | Kidane Y G, Gesesse C A. Hailemariam B N, Desta E A, Mengistu D K, Fadda C, Pè M E, Dell’Acqua M . A large nested association mapping population for breeding and quantitative trait locus mapping in Ethiopian durum wheat. Plant Biotechnol J, 2019,17:1380-1393. |
| [38] | Marla S R., Burow G, Chopra R, Hayes C, Olatoye M O, Felderhoff T, Hu Z, Raymundo R, Perumal R, Morris G P. Genetic architecture of chilling tolerance in sorghum dissected with a nested association mapping population. G3: Genes Genom Genet, 2019,9:4045-4057. |
| [39] | Gangurde S S, Wang H, Yaduru S, Pandey M K, Fountain J C, Chu Y, Isleib T, Holbrook C C, Xavier A, Culbreath A K, Ozias-Akins P, Varshney R K, Guo B. Nested-association mapping (NAM)-based genetic dissection uncovers candidate genes for seed and pod weights in peanut (Arachis hypogaea). Plant Biotechnol J, 2010,18:1457-1471. |
| [1] | 陈影,张晟瑞,王岚,王连铮,李斌,孙君明. 野生和栽培大豆种质油脂组成特点及其与演化的关系[J]. 作物学报, 2019, 45(7): 1038-1049. |
| [2] | 林婷婷,王建军,王立,陈暄,侯喜林,李英. 不结球白菜维生素C含量主基因+多基因遗传分析[J]. 作物学报, 2014, 40(10): 1733-1739. |
| [3] | 周清元,崔翠,阴涛,陈东亮,张正圣,李加纳. 甘蓝型油菜角果长度的主+多基因混合遗传模型[J]. 作物学报, 2014, 40(08): 1493-1500. |
| [4] | 贺建波,管荣展,盖钧镒. 双列杂交设计的主-微位点组遗传分析方法研究[J]. 作物学报, 2010, 36(08): 1248-1257. |
| [5] | 苏成付,赵团结,盖鈞镒. 不同统计遗传模型QTL定位方法应用效果的模拟比较 [J]. 作物学报, 2010, 36(07): 1100-1107. |
| [6] | 张启武,江建华,姚瑾,洪德林. 穞稻与粳稻恢复系C堡籽粒灌浆速率的特征及遗传分析[J]. 作物学报, 2009, 35(7): 1229-1235. |
| [7] | 刘顺湖,周瑞宝,盖钧镒. 大豆蛋白质有关性状遗传的分离分析[J]. 作物学报, 2009, 35(11): 1958-1966. |
| [8] | 李余生;朱镇;张亚东;赵凌;王才林. 水稻稻曲病抗性的主基因+多基因混合遗传模型分析[J]. 作物学报, 2008, 34(10): 1728-1733. |
| [9] | 邢光南;赵团结;盖钧镒. 大豆对豆卷叶螟Lamprosema indicata (Fabricius)抗性的遗传分析[J]. 作物学报, 2008, 34(01): 8-16. |
| [10] | 张立平;赵昌平;单福华;张风廷;叶志杰. 小麦光温敏雄性不育系BS210育性的主基因+多基因混合遗传分析[J]. 作物学报, 2007, 33(09): 1553-1557. |
| [11] | 武翠;邵国军;吕文彦;马莲菊;崔鑫福;曹萍;尹长斌;徐兴伟;何萍. 粳稻糊化温度遗传研究[J]. 作物学报, 2007, 33(06): 1041-1044. |
| [12] | 蔡长春;陈宝元;傅廷栋;涂金星. 甘蓝型油菜开花期和光周期敏感性的遗传分析[J]. 作物学报, 2007, 33(02): 345-348. |
| [13] | 易秋香;黄敬峰;王秀珍;钱翌. 玉米粗脂肪含量高光谱估算模型初探[J]. 作物学报, 2007, 33(01): 171-174. |
| [14] | 于永红;朱智伟;樊叶杨;段彬伍;庄杰云. 应用重组自交系群体检测控制水稻糙米粗蛋白和粗脂肪含量的QTL[J]. 作物学报, 2006, 32(11): 1712-1716. |
| [15] | 陈献功;刘金波;洪德林. 粳稻直立穗与弯曲穗3个杂交组合6个世代穗角和每穗颖花数的遗传分析[J]. 作物学报, 2006, 32(08): 1143-1150. |
|
||