亚麻生氰糖苷合成关键酶CYP79基因家族的鉴定及表达分析
齐燕妮, 李闻娟, 赵丽蓉, 李雯, 王利民, 谢亚萍, 赵玮, 党照, 张建平
作物学报
2023, 49 ( 3):
687-702.
DOI: 10.3724/SP.J.1006.2023.24042
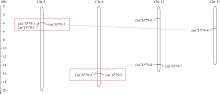
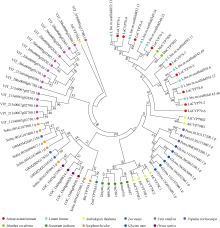
CYP79蛋白是生氰糖苷合成关键酶, 但关于亚麻CYP79基因的研究鲜有报道。本研究对包括亚麻在内的9种作物的CYP79基因家族进行了鉴定, 并分析了亚麻CYP79基因的序列特征、复制事件、共线性关系、系统进化、顺式作用元件及表达模式。结果表明, 在亚麻、白亚麻、毛果杨、木薯、芝麻、高粱、大豆、葡萄及水稻中分别鉴定到9、9、3、2、5、7、6、16和4个CYP79家族成员; 系统进化分析显示, CYP79基因的进化具有物种特异性; LuCYP79不均匀分布在4条染色体上, 具有1~3个外显子, 其启动子区含大量激素与逆境响应相关元件; 共克隆到8个亚麻CYP79基因的全长DNA序列及5个成员的全长cDNA序列; LuCYP79蛋白序列长度为282~565 aa, 等电点为5.84~9.14, 分子量为31.56~62.86 kD, 均为亲水性蛋白, 定位于内质网; 共有5对基因发生复制事件, 占全部基因的77.8%, 全部经历了强烈的纯化选择, 其中LuCYP79-1和LuCYP79-9在拟南芥和木薯中均具有同源基因。表达分析表明, LuCYP79家族成员具有组织特异性, 且各成员在不同遗传背景下表达模式不同, 其中LuCYP79-1、LuCYP79-7、LuCYP79-8和LuCYP79-9在4个品种中的表达差异显著。相关分析表明, 50 d时的LuCYP79-1/LuCYP79-7与成熟亚麻籽中生氰糖苷含量呈极显著正相关, 20 d时的LuCYP79-7/LuCYP79-8及(LuCYP79-7+LuCYP79-9)/LuCYP79-8分别与成熟亚麻籽中生氰糖苷含量呈极显著正相关, 初步推测其可能是亚麻籽生氰糖苷合成的关键基因。研究结果对进一步阐明亚麻CYP79蛋白的功能具有积极意义, 并为培育低生氰糖苷亚麻品种提供了理论参考。
| 蛋白名称Protein name | α-螺旋
Alpha helix | β-转角
Beta turn | 延伸链
Extended strand | 无规则卷曲
Random coil | | LuCYP79-9 | 120 (42.55%) | 7 (2.48%) | 35 (12.41%) | 120 (42.55%) | | LuCYP79-5 | 255 (46.03%) | 22 (3.97%) | 58 (10.47%) | 219 (39.53%) | | LuCYP79-4 | 240 (43.01%) | 32 (5.73%) | 86 (15.41%) | 200 (35.84%) | | LuCYP79-7 | 231 (43.02%) | 23 (4.28%) | 71 (13.22%) | 212 (39.48%) | | LuCYP79-8 | 177 (46.95%) | 9 (2.39%) | 48 (12.73%) | 143 (37.93%) | | LuCYP79-6 | 240 (46.07%) | 21 (4.03%) | 64 (12.28%) | 196 (37.62%) | | LuCYP79-3 | 234 (44.57%) | 19 (3.62%) | 68 (12.95%) | 204 (38.86%) | | LuCYP79-2 | 243 (44.26%) | 21 (3.83%) | 76 (13.84%) | 209 (38.07%) | | LuCYP79-1 | 247 (43.72%) | 25 ( 4.42%) | 76 (13.45%) | 217 (38.41%) |
View table in article
附表4
亚麻CYP79蛋白二级结构预测
正文中引用本图/表的段落
LuCYP79基因结构分析结果表明, 除LuCYP79-5和LuCYP79-6分别具有1个和3个外显子外, 其余基因均具有2个外显子(表1和图2)。理化性质分析结果表明, LuCYP79蛋白分子量在31.56~62.86 kD之间; 等电点在5.84~9.14之间, 只有LuCYP79-1和LuCYP79-6为酸性蛋白, 其余均为碱性蛋白; 所有LuCYP79蛋白为亲水性蛋白(附图1)。亚细胞定位结果表明, 所有LuCYP79蛋白均定位在内质网。对LuCYP79蛋白二级结构进行预测发现, α-螺旋含量最多, 其次为无规则卷曲和延伸链, β-转角含量最少(附表4)。LuCYP79蛋白三级结构预测结果与二级结构一致(附图2)。对LuCYP79蛋白进行保守基序分析发现, 9个LuCYP79蛋白共预测到14个保守基序(Motif), 其中有5个成员具有14个Motif, LuCYP79-1含有的Motif最少(图2和附图3)。Motif 1、Motif 4和Motif 8存在于所有LuCYP79成员, 而且Motif 1和Motif 4分别含有CYP79家族高度保守的血氧红素结合位点SFSTG(K/R)RGC(A/I)A和FXP(E/D)RH位点(图3)。此外, 除Motif14, 其余10个Motif存在于8个LuCYP79蛋白。
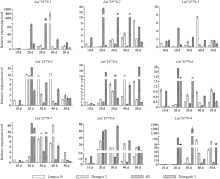
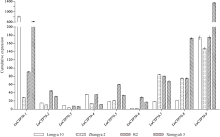
利用qRT-PCR分析9个LuCYP79家族成员在4个亚麻品种不同发育阶段种子中的表达模式发现, 各基因在不同品种中具有不同的表达模式, 如LuCYP79-1、LuCYP79-3、LuCYP79-4和LuCYP79-9在陇亚10号开花后40 d的种子中表达量最高, 但在R2中的表达量在开花后20 d达到最高, 而在匈牙利3号中, 其在开花后30 d的种子中表达量最高; LuCYP79-2在陇亚10号、张亚2号、R2及匈牙利3号中的表达量分别在开花后50 d、40 d、20 d及30 d的种子中达到最高; LuCYP79-5在4个品种种子中的表达量分别在50 d、40 d、50 d、20 d达到最高; LuCYP79-6在不同品种种子中的表达量高峰分别为10 d、50 d、20 d、20 d; LuCYP79-7的表达量高峰分别在30 d、40 d、20 d、20 d; LuCYP79-8分别在30 d、40 d、20 d、30 d的种子中表达量较高(图7)。总的来说, 各基因在张亚2号不同发育阶段种子中的表达量普遍低于其他3个品种。除表达模式外, 不同发育阶段各基因的累积表达量在不同品种中具有明显差异(图8), 其中LuCYP79-1、LuCYP79-7、LuCYP79-8和LuCYP79-9在4个品种中的差异最明显, 最高表达量是最低表达量的4倍以上, 差异最大的可达到28倍。
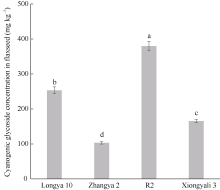
基因表达模式对深入研究基因功能具有非常重要的作用。为进一步了解LuCYP79家族各成员的功能, 本研究在不同品种、不同组织、不同发育时期对LuCYP79基因进行了表达分析。LuCYP79基因在陇亚10号、张亚2号、R2及匈牙利3号4个品种亚麻籽中具不同的表达趋势, 这可能与各品种的遗传背景相关。此外, 各基因在张亚2号中的表达量普遍较低, 这与生氰糖苷含量测定结果相一致, 张亚2号成熟籽粒中生氰糖苷含量显著低于其他3个品种。对各基因在种子不同发育阶段的累积表达量进行分析发现, 各成员的累积表达量在籽粒生氰糖苷含量具有显著差异的4个亚麻品种中差异极大, 其中LuCYP79-1、LuCYP79-7、LuCYP79-8和LuCYP79-9在4个品种中的差异达到4倍以上, 差异最大的可达到28倍。且本研究发现, 花后50 d籽粒中LuCYP79-1与LuCYP79-7基因表达量的比值, 花后20 d籽粒中LuCYP79-7与LuCYP79-8、LuCYP79-7+LuCYP79-9与LuCYP79-8基因表达量的比值分别与成熟籽粒中生氰糖苷含量呈极显著正相关(P≤0.01), 说明这几个基因可能在亚麻籽生氰糖苷合成中起关键作用。进一步分析LuCYP79-1、LuCYP79-7、LuCYP79-8和LuCYP79-9在不同组织中的表达模式发现, 这几个基因呈现出不同的组织特异性表达, 且具有不同的表达趋势, 此外, 同一个基因在不同品种中的时空表达模式也不尽相同, 说明这些基因可能在不同基因型亚麻品种生氰糖苷合成或是其他生理过程中发挥不同的作用, 但各成员具体的功能还需深入探究。
本文的其它图/表
|