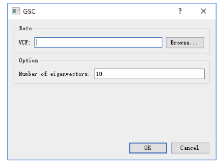
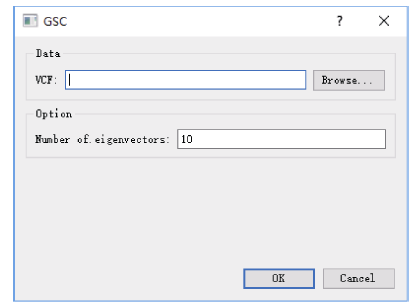
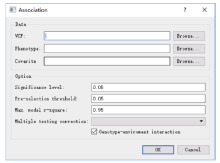
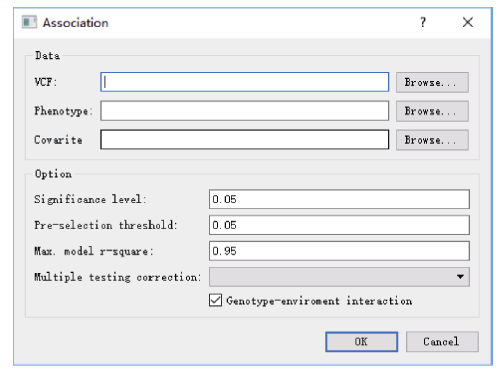
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2018, Vol. 44 ›› Issue (9): 1274-1289.doi: 10.3724/SP.J.1006.2018.01274
• RESEARCH PAPERS • Previous Articles Next Articles
Jian-Bo HE( ),Fang-Dong LIU,Guang-Nan XING,Wu-Bin WANG,Tuan-Jie ZHAO,Rong-Zhan GUAN,Jun-Yi GAI()
),Fang-Dong LIU,Guang-Nan XING,Wu-Bin WANG,Tuan-Jie ZHAO,Rong-Zhan GUAN,Jun-Yi GAI()
| [1] | Visscher P M, Wray N R, Zhang Q, Sklar P , McCarthy M I, Brown M A, Yang J . 10 Years of GWAS discovery: biology, function, and translation. Am J Hum Genet, 2017,101:5-22 |
| [2] |
Pritchard J K, Stephens M, Rosenberg N A, Donnelly P . Association mapping in structured populations. Am J Hum Genet, 2000,67:170-181
doi: 10.1086/302959 pmid: 10827107 |
| [3] | Price A L, Patterson N J, Plenge R M, Weinblatt M E, Shadick N A, Reich D . Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet, 2006,38:904-909 |
| [4] | Yu J, Pressoir G, Briggs W H, Vroh Bi I, Yamasaki M, Doebley J F , McMullen M D, Gaut B S, Nielsen D M, Holland J B, Kresovich S, Buckler E S. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat Genet, 2006,38:203-208 |
| [5] |
Kang H M, Zaitlen N A, Wade C M, Kirby A, Heckerman D, Daly M J, Eskin E . Efficient control of population structure in model organism association mapping. Genetics, 2008,178:1709-1723
doi: 10.1534/genetics.107.080101 pmid: 18385116 |
| [6] |
Kang H M, Sul J H, Service S K, Zaitlen N A, Kong S Y, Freimer N B, Sabatti C, Eskin E . Variance component model to account for sample structure in genome-wide association studies. Nat Genet, 2010,42:348-354
doi: 10.1038/ng.548 pmid: 20208533 |
| [7] |
Zhang Z, Ersoz E, Lai C Q, Todhunter R J, Tiwari H K, Gore M A, Bradbury P J, Yu J, Arnett D K, Ordovas J M, Buckler E S . Mixed linear model approach adapted for genome-wide association studies. Nat Genet, 2010,42:355-360
doi: 10.1038/ng.546 pmid: 20208535 |
| [8] |
Pritchard J K, Stephens M, Donnelly P . Inference of population structure using multilocus genotype data. Genetics, 2000,155:945-959
pmid: 10835412 |
| [9] | Alexander D H, Novembre J, Lange K . Fast model-based estimation of ancestry in unrelated individuals. Genome Res, 2009,19:1655-1664 |
| [10] | Atwell S, Huang Y S, Vilhjalmsson B J, Willems G, Horton M, Li Y, Meng D, Platt A, Tarone A M, Hu T T, Jiang R, Muliyati N W, Zhang X, Amer M A, Baxter I, Brachi B, Chory J, Dean C , Debieu M, de Meaux J, Ecker J R, Faure N, Kniskern J M, Jones J D, Michael T, Nemri A, Roux F, Salt D E, Tang C, Todesco M, Traw M B, Weigel D, Marjoram P, Borevitz J O, Bergelson J, Nordborg M . Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature, 2010,465:627-631 |
| [11] |
Huang X, Wei X, Sang T, Zhao Q, Feng Q, Zhao Y, Li C, Zhu C, Lu T, Zhang Z, Li M, Fan D, Guo Y, Wang A, Wang L, Deng L, Li W, Lu Y, Weng Q, Liu K, Huang T, Zhou T, Jing Y, Li W, Lin Z, Buckler E S, Qian Q, Zhang Q F, Li J, Han B . Genome-wide association studies of 14 agronomic traits in rice landraces. Nat Genet, 2010,42:961-967
doi: 10.1038/ng.695 pmid: 20972439 |
| [12] | Li H, Peng Z, Yang X, Wang W, Fu J, Wang J, Han Y, Chai Y, Guo T, Yang N, Liu J, Warburton M L, Cheng Y, Hao X, Zhang P, Zhao J, Liu Y, Wang G, Li J, Yan J . Genome-wide association study dissects the genetic architecture of oil biosynthesis in maize kernels. Nat Genet, 2013,45:43-50 |
| [13] | Fang C, Ma Y M, Wu S W, Liu Z, Wang Z, Yang R, Hu G H, Zhou Z K, Yu H, Zhang M, Pan Y, Zhou G A, Ren H X, Du W Q, Yan H R, Wang Y P, Han D Z, Shen Y T, Liu S L, Liu T F, Zhang J X, Qin H, Yuan J, Yuan X H, Kong F J, Liu B H, Li J Y, Zhang Z W, Wang G D, Zhu B G, Tian Z X . Genome-wide association studies dissect the genetic networks underlying agronomical traits in soybean. Genome Biol, 2017,18:161 |
| [14] | Peleman J D , van der Voort J R. Breeding by design. Trends Plant Sci, 2003,8:330-334 |
| [15] |
Huang X, Wei X, Sang T, Zhao Q, Feng Q, Zhao Y, Li C, Zhu C, Lu T, Zhang Z, Li M, Fan D, Guo Y, Wang A, Wang L, Deng L, Li W, Lu Y, Weng Q, Liu K, Huang T, Zhou T, Jing Y, Li W, Lin Z, Buckler E S, Qian Q, Zhang Q F, Li J, Han B . Genome-wide association studies of 14 agronomic traits in rice landraces. Nat Genet, 2010,42:961-967
doi: 10.1038/ng.695 pmid: 20972439 |
| [16] | Zhao K, Tung C W, Eizenga G C, Wright M H, Ali M L, Price A H, Norton G J, Islam M R, Reynolds A, Mezey J , McClung A M, Bustamante C D, McCouch S R . Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat Commun, 2011,2:467 |
| [17] |
Zeng Z B . Precision mapping of quantitative trait loci. Genetics, 1994,136:1457-1468
doi: 10.1007/s00122-012-2032-2. pmid: 8013918 |
| [18] | He J, Meng S, Zhao T, Xing G, Yang S, Li Y, Guan R, Lu J, Wang Y, Xia Q, Yang B, Gai J . An innovative procedure of genome-wide association analysis fits studies on germplasm population and plant breeding. Theor Appl Genet, 2017,130:2327-2343 |
| [19] | Zhang Y, He J, Wang Y, Xing G, Zhao J, Li Y, Yang S, Palmer R G, Zhao T, Gai J . Establishment of a 100-seed weight quantitative trait locus-allele matrix of the germplasm population for optimal recombination design in soybean breeding programmes. J Exp Bot, 2015,66:6311-6325 |
| [20] | Meng S, He J, Zhao T, Xing G, Li Y, Yang S, Lu J, Wang Y, Gai J . Detecting the QTL-allele system of seed isoflavone content in Chinese soybean landrace population for optimal cross design and gene system exploration. Theor Appl Genet, 2016,129:1557-1576 |
| [21] | Li S, Cao Y, He J, Zhao T, Gai J . Detecting the QTL-allele system conferring flowering date in a nested association mapping population of soybean using a novel procedure. Theor Appl Genet, 2017,130:2297-2314 |
| [22] | Gabriel S B, Schaffner S F, Nguyen H, Moore J M, Roy J, Blumenstiel B, Higgins J , DeFelice M, Lochner A, Faggart M, Liu-Cordero S N, Rotimi C, Adeyemo A, Cooper R, Ward R, Lander E S, Daly M J, Altshuler D . The structure of haplotype blocks in the human genome. Science, 2002,296:2225-2229 |
| [23] | Patterson N, Price A L, Reich D . Population structure and eigenanalysis. PLoS Genet, 2006,2:e190 |
| [24] | Price A L, Patterson N J, Plenge R M, Weinblatt M E, Shadick N A, Reich D . Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet, 2006,38:904-909 |
| [25] |
VanRaden P M . Efficient methods to compute genomic predictions. J Dairy Sci, 2008,91:4414-4423
doi: 10.3168/jds.2007-0980 pmid: 18946147 |
| [26] | Scheet P, Stephens M . A fast and flexible statistical model for large-scale population genotype data: applications to inferring missing genotypes and haplotypic phase. Am J Hum Genet, 2006,78:629-644 |
| [27] |
Buckler E S, Holland J B, Bradbury P J, Acharya C B, Brown P J, Browne C, Ersoz E, Flint-Garcia S, Garcia A, Glaubitz J C, Goodman M M, Harjes C, Guill K, Kroon D E, Larsson S, Lepak N K, Li H, Mitchell S E, Pressoir G, Peiffer J A, Rosas M O, Rocheford T R, Romay M C, Romero S, Salvo S , Sanchez Villeda H, da Silva H S, Sun Q, Tian F, Upadyayula N, Ware D, Yates H, Yu J, Zhang Z, Kresovich S, McMullen M D. The genetic architecture of maize flowering time. Science, 2009,325:714-718
doi: 10.1126/science.1174276 pmid: 19661422 |
| [28] |
Meuwissen T H, Hayes B J, Goddard M E . Prediction of total genetic value using genome-wide dense marker maps. Genetics, 2001,157:1819-1829
doi: 10.1017/S0016672301004931 pmid: 11290733 |
| [1] | FENG Jian-Ying,WEN Yang-Jun,ZHANG Jin,ZHANG Yuan-Ming. Advances on Methodologies for Genome-wide Association Studies in Plants [J]. Acta Agron Sin, 2016, 42(07): 945-956. |
|
||