欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (10): 2546-2559.doi: 10.3724/SP.J.1006.2022.14148
唐映红1,3,4( ), 刘芳2, 陈建荣2,*(), 毛凯权2, 李辉1,3, 万海清1
), 刘芳2, 陈建荣2,*(), 毛凯权2, 李辉1,3, 万海清1
TANG Ying-Hong1,3,4(), LIU Fang2, CHEN Jian-Rong2,*(), MAO Kai-Quan2, LI Hui1,3, WAN Hai-Qing1
摘要:
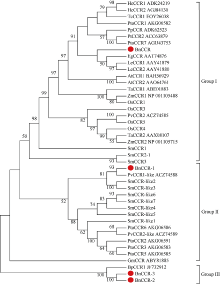
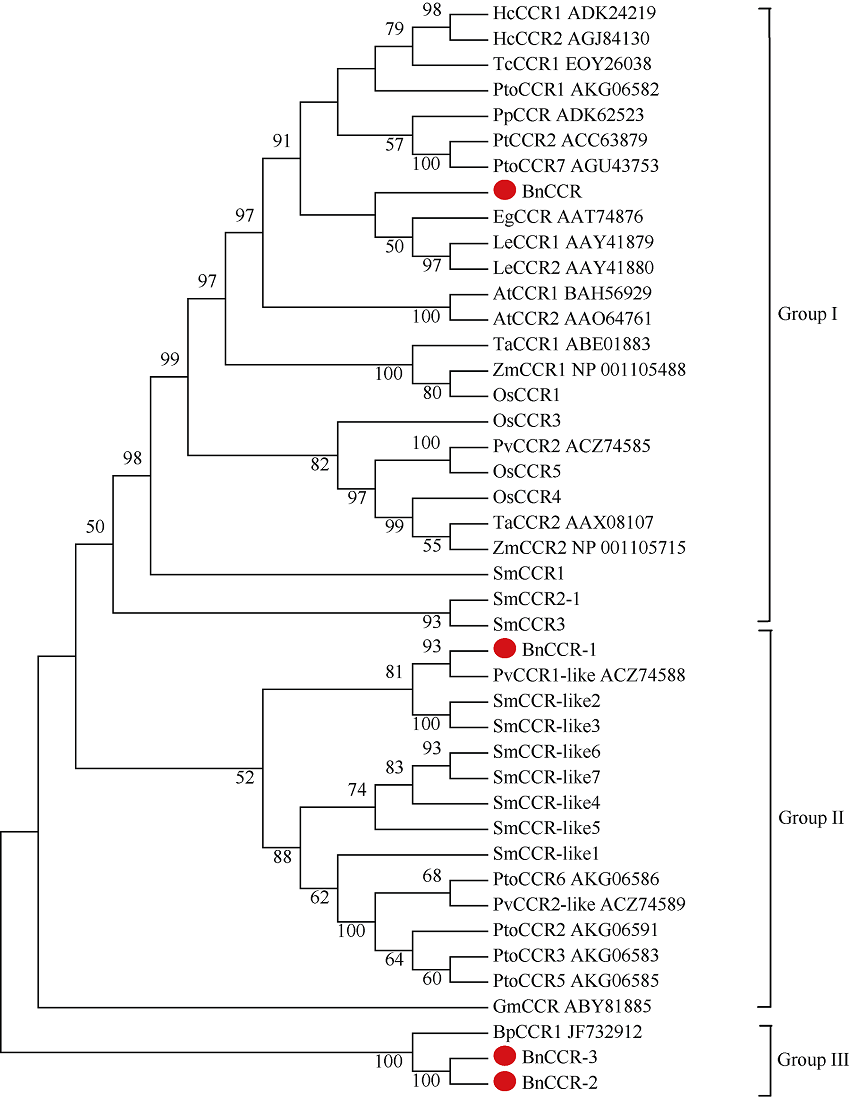
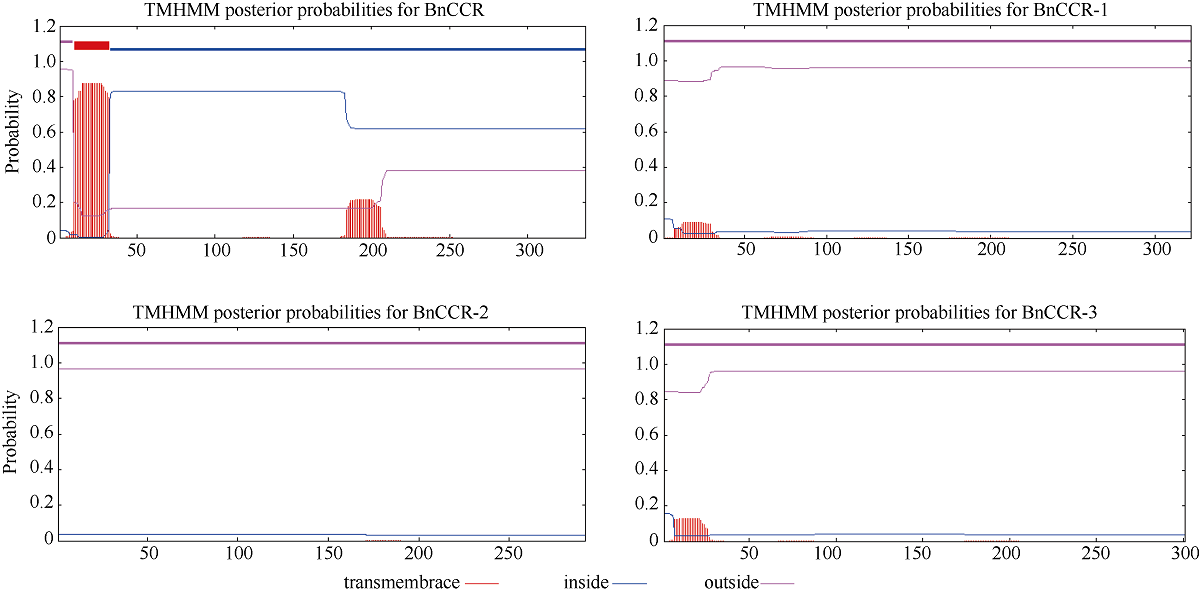
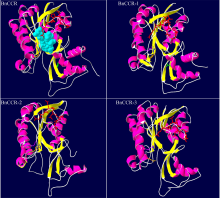
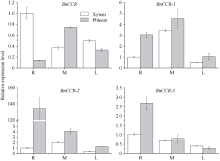
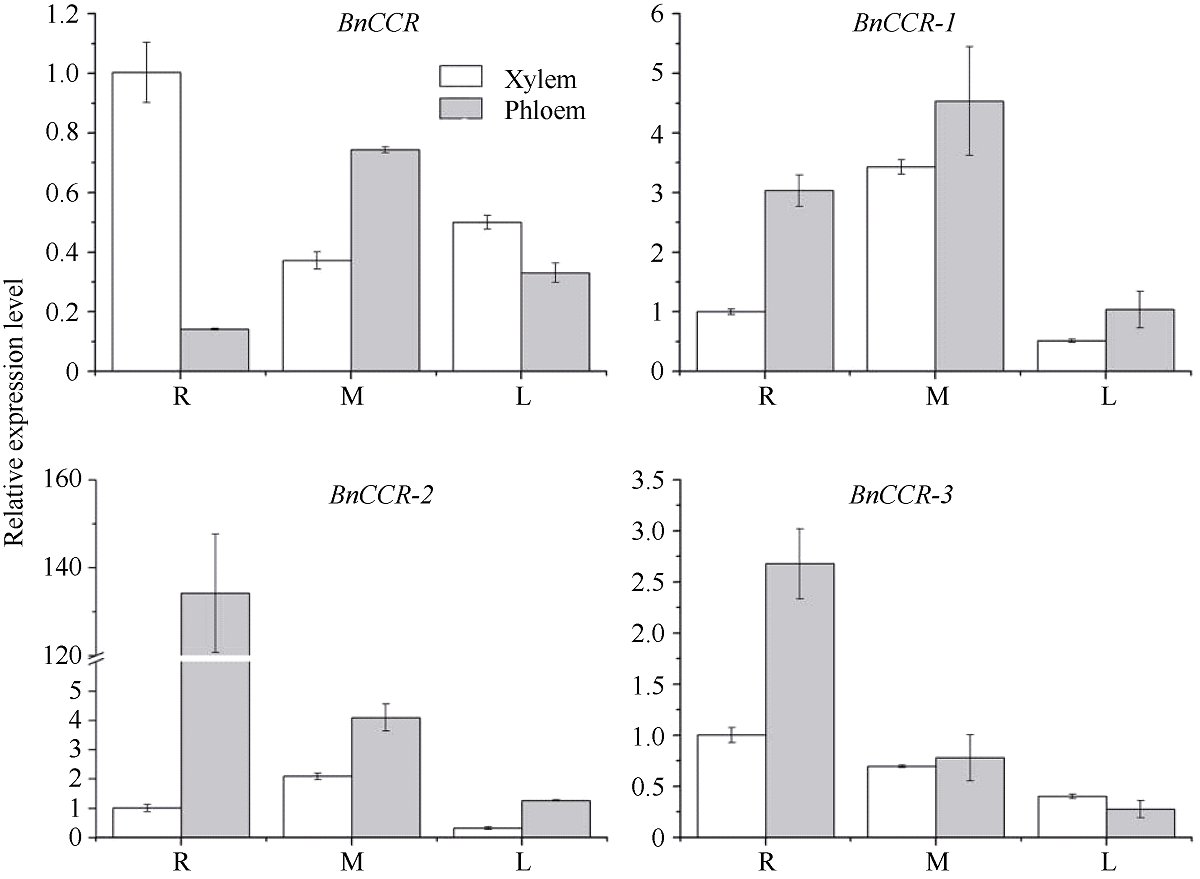
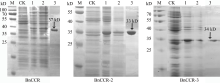
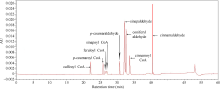
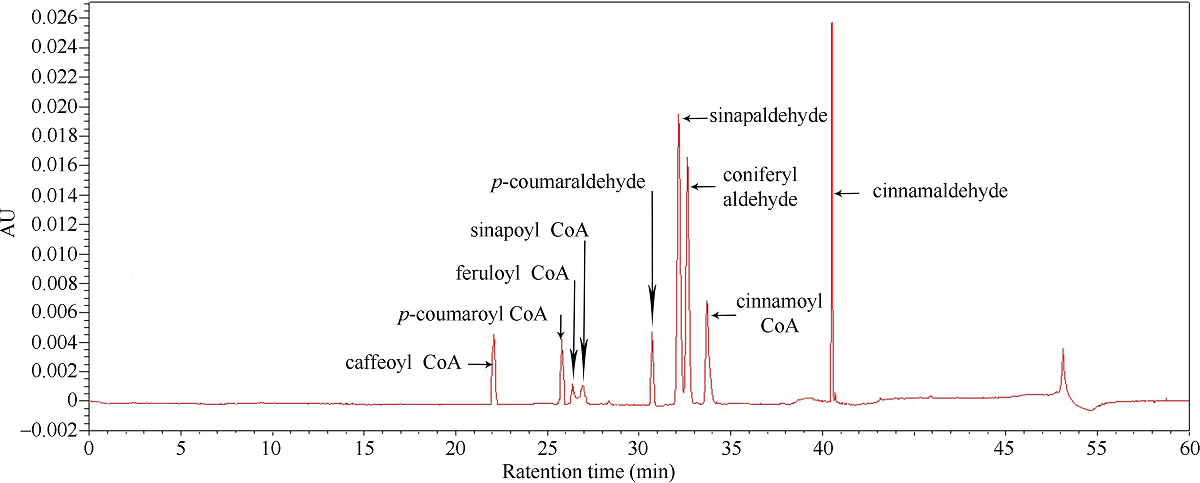
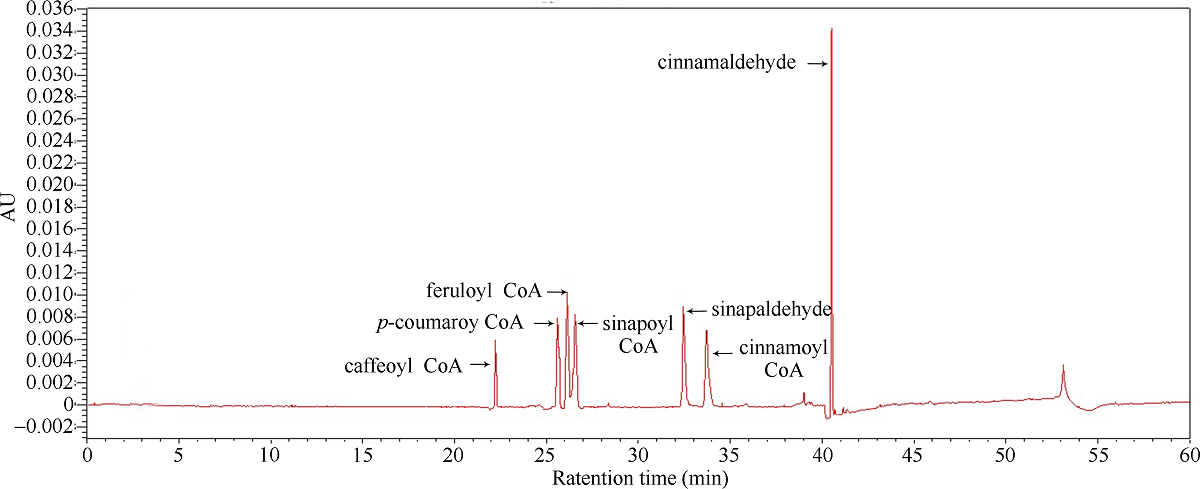
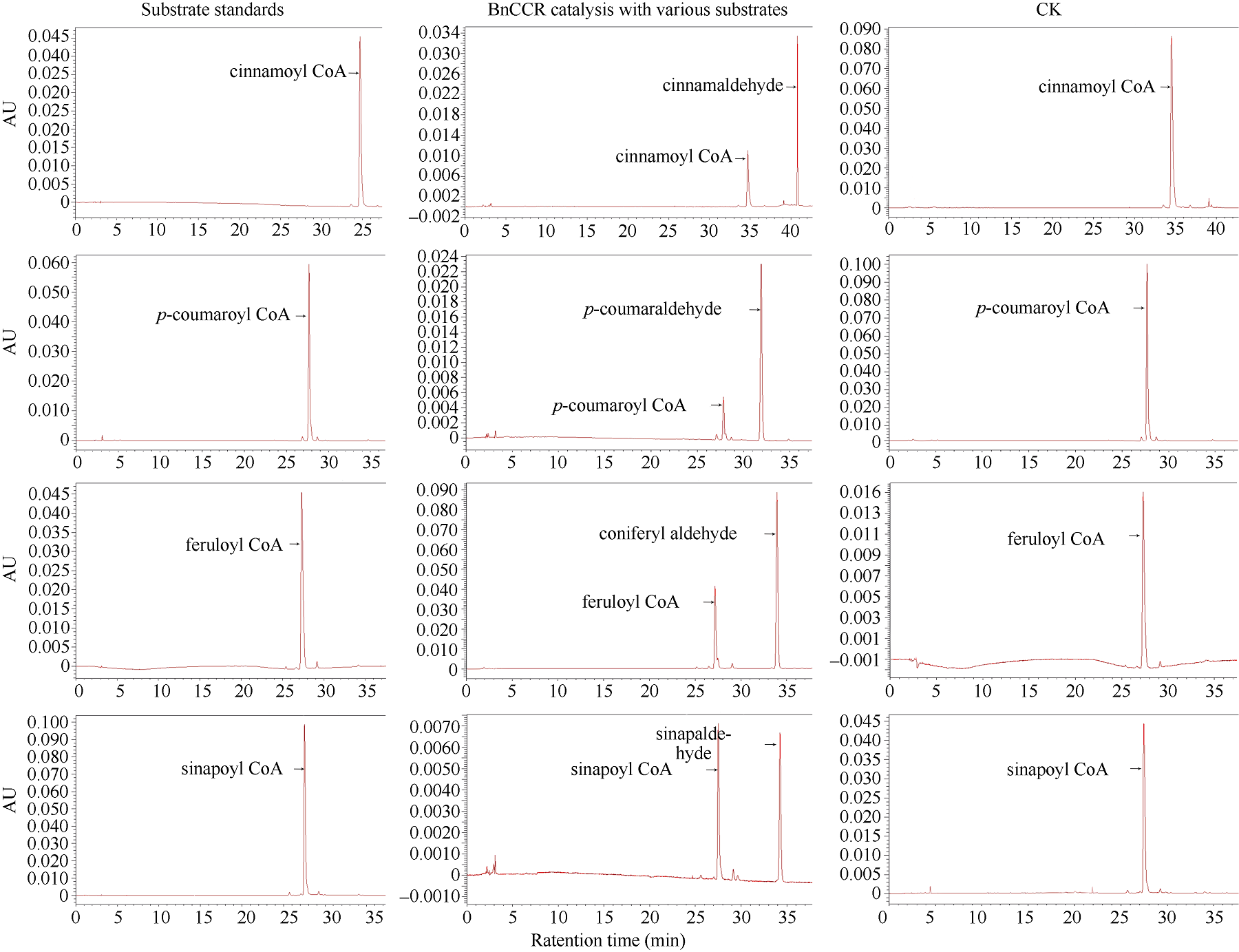
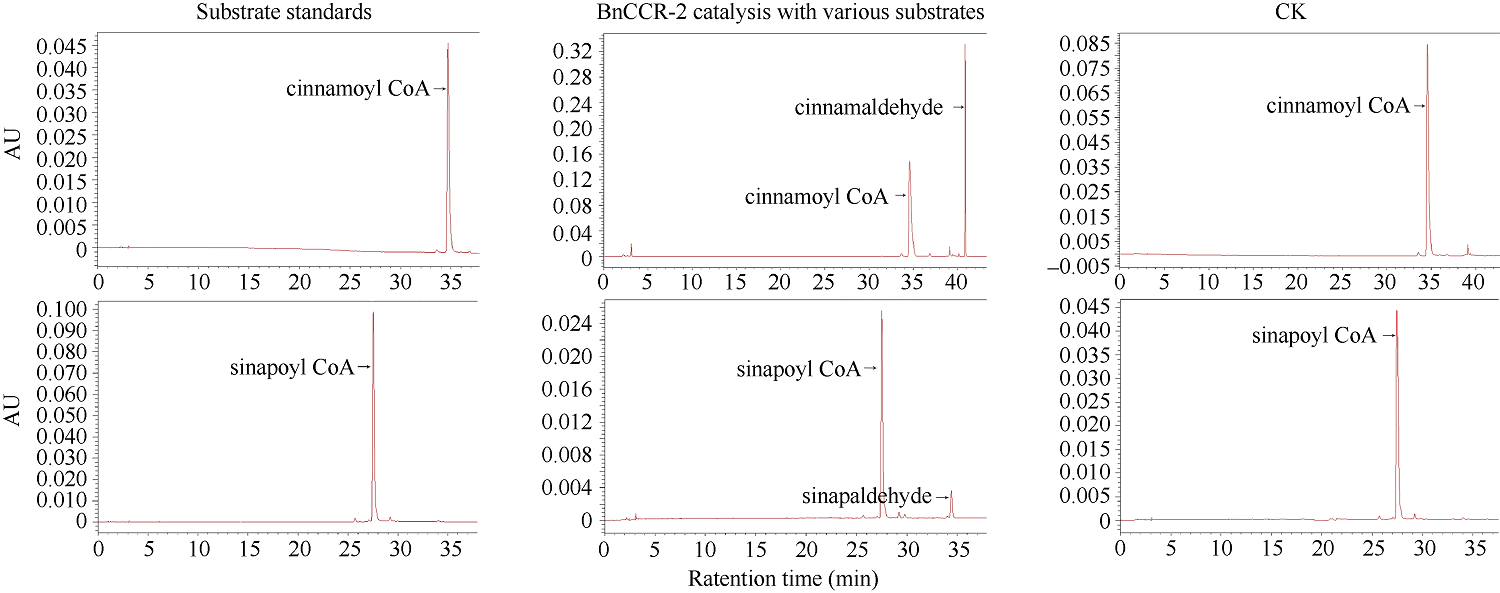
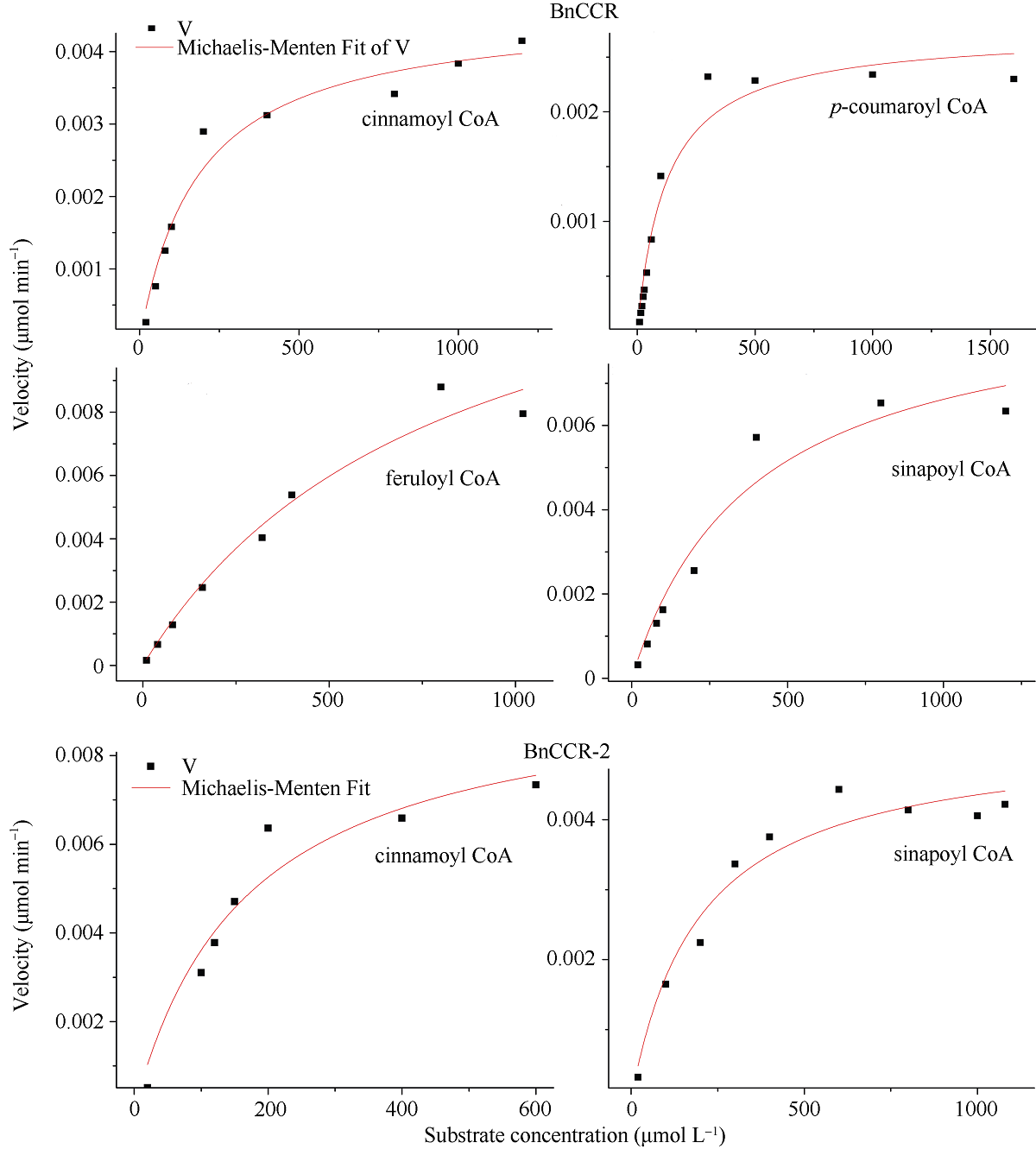
肉桂酰辅酶A还原酶(CCR)是木质素生物合成的关键酶, 在植物中多以基因家族形式存在。本研究通过苎麻转录组信息, 筛选并克隆了4个苎麻CCR基因家族成员(BnCCR、BnCCR-1、BnCCR-2和BnCCR-3)。序列分析结果发现4个BnCCRs可以分为3类, BnCCR属于bona fide CCR类群, BnCCR-1为CCR类似蛋白, 而BnCCR-2和BnCCR-3序列在NADPH结合功能域、催化三联体以及CCR底物结合位点上均与bona fide CCR都有差异, 形成了苎麻中一个新的CCR类群。跨膜结构分析显示, BnCCR含有1个跨膜螺旋区, 其他3个BnCCRse无跨膜螺旋区。结构分析显示, 4个BnCCRs的二级和三级结构存在差异, BnCCR-2同源建模模板不同于其他3个BnCCRs。时空表达谱分析发现, BnCCR在快速生长期的木质部具有较高表达水平, 而BnCCR-2在快速生长期的韧皮部具有非常高的表达水平。体外酶活测定进一步研究发现, BnCCR具有典型的bona fide CCR催化活性和底物适应性, 而BnCCR-2的表现不同于典型CCR的底物适应性, 对肉桂酰CoA和芥子酰CoA表现出结合特异性。因此, 苎麻中BnCCR参与木质素的生物合成, 而BnCCR-2是具有不同于典型CCR结构、组织表达和体外生化功能的一个新类群, BnCCR-2可能不参与或并不仅仅参与木质素的生物合成, 这为研究CCR蛋白家族的进化提供了新的参考。
| [1] |
Zhao Q. Lignification: flexibility, biosynthesis and regulation. Trends Plant Sci, 2016, 21: 713-721.
doi: 10.1016/j.tplants.2016.04.006 |
| [2] | 郭亚玉, 许会敏, 赵媛媛, 吴鸿洋, 林金星. 植物木质化过程及其调控的研究进展. 中国科学: 生命科学, 2020, 50: 111-122. |
|
Guo Y Y, Xu H M, Zhao Y Y, Wu H X, Lin J X. Plant lignification and its regulation. Sci Sin (Vitae), 2020, 50: 111-122 (in Chinese with English abatract).
doi: 10.2298/SOS1801111K |
|
| [3] |
Day A, Ruel K, Neutelings G, Cronier D, David H, Hawkins S, Chabbert B. Lignification in the flax stem: evidence for an unusual lignin in bast fibers. Planta, 2005, 222: 234-245.
doi: 10.1007/s00425-005-1537-1 |
| [4] | 袁有美, 唐映红, 刘芳, 陈建荣. 免耕种植水土保持用苎麻木质纤维主要成分分析. 中国农学通报, 2015, 31(31): 242-246. |
| Yuan Y M, Tang Y H, Liu F, Chen J R. Analysis of fiber main ingredients in no-till planting ramie. Chin Agric Sci Bull, 2015, 31(31): 242-246. (in Chinese with English abstract) | |
| [5] |
徐益, 张力岚, 祁建民, 张列梅, 张立武. 主要麻类作物基因组学与遗传改良: 现状与展望. 作物学报, 2021, 47: 997-1019.
doi: 10.3724/SP.J.1006.2021.04121 |
| Xu Y, Zhang L L, Qi J M, Zhang L M, Zhang L W. Genomics and genetic improvement in main bast fiber crops: advances and perspectives. Acta Agron Sin, 2021, 47: 997-1019. (in Chinese with English abstract) | |
| [6] |
Sonawane P, Vishwakarma R K, Khan B M. Biochemical characterization of recombinant cinnamoyl CoA reductase 1 (Ll- CCRH1) from Leucaena leucocephala. Int J Biol Macromol, 2013, 58: 154-159.
doi: 10.1016/j.ijbiomac.2013.03.050 |
| [7] |
Barakat A, Yassin N B M, Park J S, Choi A, Herr J, Carlson J E. Comparative and phylogenomic analyses of cinnamoyl-CoA reductase and cinnamoyl-CoA-reductase-like gene family in land plants. Plant Sci, 2011, 181: 249-257.
doi: 10.1016/j.plantsci.2011.05.012 |
| [8] | Labeeuw L, Martone P T, Boucher Y, Case R J. Ancient origin of the biosynthesis of lignin precursors. Biol Direct, 2015, 10: 23. |
| [9] |
Escamilla-Trevino L L, Shen H, Uppalapati S R, Ray T, Tang Y, Hernandez T, Yin Y, Xu Y, Dixon R A. Switchgrass (Panicum virgatum) possesses a divergent family of cinnamoyl CoA reductases with distinct biochemical properties. New Phytol, 2010, 185: 143-155.
doi: 10.1111/j.1469-8137.2009.03018.x pmid: 19761442 |
| [10] |
Zhou R, Jackson L, Shadle G, Nakashima J, Temple S, Chen F, Dixon R A. Distinct cinnamoyl CoA reductases involved in parallel routes to lignin in Medicago truncatula. Proc Natl Acad Sci USA, 2010, 107: 17803-17808.
doi: 10.1073/pnas.1012900107 |
| [11] |
Chao N, Li N, Qi Q, Li S, Lyu T, Jiang X N, Gai Y. Characterization of the cinnamoyl-CoA reductase (CCR) gene family in Populus tomentosa reveals the enzymatic active sites and evolution of CCR. Planta, 2017, 245: 61-75.
doi: 10.1007/s00425-016-2591-6 pmid: 27580618 |
| [12] |
Baltas M, Lapeyre C, Bedos-Belval F, Maturano M, Saint-Aguet P, Roussel L, Duran H, Grima-Pettenati J. Kinetic and inhibition studies of cinnamoyl-CoA reductase 1 from Arabidopsis thaliana. Plant Physiol Biochem, 2005, 43: 746-753.
doi: 10.1016/j.plaphy.2005.06.003 |
| [13] |
Sonawane P, Patel K, Vishwakarma R K, Srivastava S, Singh S, Gaikwad S, Khan B M. Probing the active site of cinnamoyl CoA reductase 1 (Ll-CCRH1) from Leucaena leucocephala. Int J Biol Macromol, 2013, 60: 33-38.
doi: 10.1016/j.ijbiomac.2013.05.005 |
| [14] |
Bomati E K, Noel J P. Structural and kinetic basis for substrate selectivity in Populus tremuloides sinapyl alcohol dehydrogenase. Plant Cell, 2005, 17: 1598-1611.
pmid: 15829607 |
| [15] |
Ma Q H. Characterization of a cinnamoyl-CoA reductase that is associated with stem development in wheat. J Exp Bot, 2007, 58: 2011-2021.
doi: 10.1093/jxb/erm064 |
| [16] |
Lauvergeat V, Lacomme C, Lacombe E, Lasserre E, Roby D, Grima-Pettenati J. Two cinnamoyl-CoA reductase (CCR) genes from Arabidopsis thaliana are differentially expressed during development and in response to infection with pathogenic bacteria. Phytochemistry, 2001, 57: 1187-1195.
pmid: 11430991 |
| [17] |
Goffner D, Campbell M M, Campargue C, Clastre M, Borderies G, Boudet A, Boudet A M. Purification and characterization of cinnamoyl-coenzyme A: NADP oxidoreductase in Eucalyptus gunnii. Plant Physiol, 1994, 106: 625-632.
pmid: 12232355 |
| [18] |
Pinçon G, Maury S, Hoffmann L, Geoffroy P, Lapierre C, Pollet B, Legrand M. Repression of O-methyltransferase genes in transgenic tobacco affects lignin synthesis and plant growth. Phytochemistry, 2001, 57: 1167-1176.
pmid: 11430989 |
| [19] | 李雪平, 彭镇华, 高志民, 胡陶. 抑制COMT基因表达对转基因烟草木质素合成的影响. 分子植物育种, 2012, 10: 689-692. |
| Li X P, Peng Z H, Gao Z M, Hu T. The Effects of depressing expression of COMT on lignin synthesis of transgenic tobacco. Mol Plant Breed, 2012, 10: 689-692. (in Chinese with English abstract) | |
| [20] | Li Y, Kim J I, Pysh L, Chapple C. Four isoforms of Arabidopsis thaliana 4-coumarate: CoA ligase (4CL) have overlapping yet distinct roles in phenylpropanoid metabolism. Plant Physiol, 2015, 169: 2409-2421. |
| [21] | Wang Q, Dai X R, Pang H Y, Cheng Y X, Huang X, Li H, Yan X J, Lu F C, Wei H R, Sederoff R R, Li Q Z. BEL1-like homeodomain protein BLH6a is a negative regulator of CAl5H2 in sinapyl alcohol monolignol biosynthesis in poplar. Front Plant Sci, 2021, 12: 1-14. |
| [22] | Chen J R, Liu F, Tang Y H, Yuan Y M, Guo Q Q. Transcriptome sequencing and profiling of expressed genes in phloem and xylem of ramie (Boehmeria nivea L. Gaud). PLoS One, 2014, 10: e110623. |
| [23] |
唐映红, 陈建荣, 刘芳, 袁有美, 郭清泉, 昌洪涛. 苎麻肉桂酰辅酶A还原酶基因cDNA序列的克隆与分析. 作物学报, 2015, 41: 1324-1332.
doi: 10.3724/SP.J.1006.2015.01324 |
|
Tang Y H, Chen J R, Liu F, Yuan Y M, Guo Q Q, Chang H T. cDNA Cloning and analysis of Cinnamoyl-CoA reductase gene from Boehmeria nivea. Acta Agron Sin, 2015, 41: 1324-1332. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2015.01324 |
|
| [24] |
Tang Y H, Liu F, Mao K Q, Xing H C, Chen J R, Guo Q Q. Cloning and characterization of the key 4-coumarate CoA ligase genes in Boehmeria nivea. South Afr J Bot, 2018, 116: 123-130.
doi: 10.1016/j.sajb.2018.02.398 |
| [25] | Tang Y H, Liu F, Xing H C, Mao K Q, Chen G, Guo Q Q, Chen J R. Correlation analysis of lignin accumulation and expression of key genes involved in lignin biosynthesis of ramie (Boehmeria nivea). Genes, 2019, 10: 389. |
| [26] |
Luan M B, Jian J B, Chen P, Chen J H, Chen J H, Gao Q, Gao G, Zhou J H, Chen K M, Guang X M, Chen J K, Zhang Q Q, Wang X F, Fang L, Sun Z M, Bai M Z, Fang X D, Zhao S C, Xiong H P, Yu C M, Zhu A G. Draft genome sequence of ramie, Boehmeria nivea (L.) Gaudich. Mol Ecol Resour, 2018, 18: 639-645.
doi: 10.1111/1755-0998.12766 |
| [27] |
Chao N, Li S, Li N, Qi Q, Jiang W T, Jiang X N, Gai Y. Two distinct cinnamoyl-CoA reductases in Selaginella moellendorffii offer insight into the divergence of CCRs in plants. Planta, 2017, 246: 33-43.
doi: 10.1007/s00425-017-2678-8 pmid: 28321576 |
| [28] |
Pan H, Zhou R, Louie G V, Mühlemann J K, Bomati E K, Bowman M E, Dudareva N, Dixon R A, Noel J P, Wang X Q. Structural studies of cinnamoyl-CoA reductase and cinnamyl-alcohol dehydrogenase, key enzymes of monolignol biosynthesis. Plant Cell, 2014, 26: 3709-3727.
doi: 10.1105/tpc.114.127399 |
| [29] |
Sattler S A, Walker A M, Vermerris W, Sattler S E, Kang C. Structural and biochemical characterization of Cinnamoyl-CoA reductases. Plant Physiol, 2017, 173: 1031-1044.
doi: 10.1104/pp.16.01671 pmid: 27956488 |
| [30] |
Chao N, Jiang W T, Wang X C, Jiang X N, Gai Y. Novel motif is capable of determining CCR and CCR-like proteins based on the divergence of CCRs in plants. Tree Physiol, 2019, 39: 2019-2026.
doi: 10.1093/treephys/tpz098 pmid: 31748812 |
| [31] |
Jones L, Ennos A R, Turner S R. Cloning and characterization of irregular xylem4 (irx4) a severely lignin-deficient mutant of Arabidopsis. Plant J, 2001, 26: 205-216.
pmid: 11389761 |
| [32] | 宋恩慧, 蔡诚, 魏国, 高慧, 项艳. RNA干涉培育低木质素杨树. 安徽林业科技, 2012, 38(2): 58-62. |
| Song E H, Cai C, Wei G, Gao H, Xiang Y. Research on RNA- interference breeding of low- lignin poplar varieties. Anhui For Sci Technol, 2012, 38(2): 58-62 (in Chinese with English abatract). | |
| [33] |
Wadenbäck J, von Arnold S, Egertsdotter U, Walter M H, Grima-Pettenati J, Goffner D, Gellerstedt G, Gullion T, Clapham D. Lignin biosynthesis in transgenic Norway spruce plants harboring an antisense construct for cinnamoyl CoA reductase (CCR). Transgenic Res, 2008, 17: 379-392.
pmid: 17610137 |
| [34] |
Kawasaki T, Koita H, Nakatsubo T, Hasegawa K, Wakabayashi K, Takahashi H, Umemura K, Umezawa T, Shimamoto K. Cinnamoyl-CoA reductase, a key enzyme in lignin biosynthesis, is an effector of small GTPase Rac in defense signaling in rice. Proc Natl Acad Sci USA, 2006, 103: 230-235.
doi: 10.1073/pnas.0509875103 |
| [1] | 李富, 王延周, 严理, 朱四元, 刘头明. 苎麻茎皮环状RNA表达谱分析[J]. 作物学报, 2021, 47(6): 1020-1030. |
| [2] | 付虹雨, 崔国贤, 李绪孟, 佘玮, 崔丹丹, 赵亮, 苏小惠, 王继龙, 曹晓兰, 刘婕仪, 刘皖慧, 王昕惠. 基于无人机遥感图像的苎麻产量估测研究[J]. 作物学报, 2020, 46(9): 1448-1455. |
| [3] | 唐映红, 陈建荣, 刘芳, 袁有美, 郭清泉, 昌洪涛. 苎麻肉桂酰辅酶A还原酶基因cDNA序列的克隆与分析[J]. 作物学报, 2015, 41(09): 1324-1332. |
| [4] | 刘昱翔,陈建荣,彭彦,黄妤,赵燕,黄丽华,郭清泉,张学文. 两种苎麻纤维素合酶基因cDNA序列的克隆及表达[J]. 作物学报, 2014, 40(11): 1925-1935. |
| [5] | 马田田,彭琦,陈松,张洁夫. 核盘菌诱导下甘蓝型油菜防御相关基因表达差异分析[J]. 作物学报, 2014, 40(03): 416-423. |
| [6] | 周精华,余伟林,邢虎成,揭雨成,钟英丽,敬礼恒. 苎麻ACC合酶基因(BnACS1)的克隆和表达分析[J]. 作物学报, 2012, 38(12): 2306-2311. |
| [7] | 夏家平,郭会君,谢永盾,赵林姝,古佳玉,赵世荣,李军辉,刘录祥. 小麦叶绿素缺失突变体Mt135的叶绿体基因差异表达分析[J]. 作物学报, 2012, 38(11): 2122-2130. |
| [8] | 邹自征,陈建华,栾明宝,郭劲霞,王超,王晓飞,许英,孙志民. 应用RSAP、SRAP和SSR分析苎麻种质亲缘关系[J]. 作物学报, 2012, 38(05): 840-847. |
| [9] | 周精华, 邢虎成, 揭雨成, 钟英丽, 朱守晶, 蒋杰, 王亮. 苎麻Δ1-吡咯啉-5-羧酸合成酶(P5CS)基因的克隆和表达分析[J]. 作物学报, 2012, 38(03): 549-555. |
| [10] | 佘玮, 揭雨成, 邢虎成, 鲁雁伟, 黄明, 康万利, 王栋. 苎麻耐镉品种差异及其筛选指标分析[J]. 作物学报, 2011, 37(02): 348-354. |
| [11] | 栾明宝, 陈建华, 许英, 王晓飞, 孙志民. 苎麻核心种质构建方法[J]. 作物学报, 2010, 36(12): 2099-2106. |
| [12] | 马雄风,喻春明,唐守伟,朱爱国,王延周,朱四元,刘建新,熊和平. 苎麻Actinl基因克隆及其在韧皮部纤维不同发育阶段的表达[J]. 作物学报, 2010, 36(1): 101-108. |
| [13] | 马雄风,喻春明,唐守伟,郭三堆,张锐,王延周,朱爱国,朱四元,熊和平. 根癌农杆菌介导的转双价抗虫基因(CryIA+CpTI)苎麻[J]. 作物学报, 2010, 36(05): 788-793. |
| [14] | 符家平,汪波,刘立军,杨金雨,王绪霞,邢秀龙,彭定祥. 根癌农杆菌介导转Bt基因苎麻的获得及其抗虫鉴定[J]. 作物学报, 2009, 35(10): 1771-1777. |
| [15] | 廖亮,李同建,刘中来,邓辉胜,徐玲玲,潘其辉,赖占均,石庆华. 基于细胞学和DNA序列的苎麻与其野生近缘类群系统关系研究[J]. 作物学报, 2009, 35(10): 1778-1790. |
|