o2突变引起糯玉米籽粒淀粉积累差异研究
韩洁楠, 张泽, 刘晓丽, 李冉, 上官小川, 周婷芳, 潘越, 郝转芳, 翁建峰, 雍洪军, 周志强, 徐晶宇, 李新海, 李明顺
作物学报
2024, 50 ( 5):
1207-1222.
DOI: 10.3724/SP.J.1006.2024.33046
糯玉米是主要鲜食玉米类型, opaque2 (o2)基因导入可增加籽粒赖氨酸含量, 但同时引起籽粒皱缩、淀粉含量下降等, 限制了其育种应用。为发掘优良糯玉米受体, 以籽粒饱满圆型o2近等基因系(o2-NIL)糯2/wx1wx1o2o2和皱缩型黄糯2/wx1wx1o2o2为研究材料, 通过对鲜食期、成熟期的百粒重和籽粒成分测定, 发现淀粉和可溶性糖含量不同可能是导致2份糯玉米o2-NILs表型差异的主要原因。利用实时荧光定量PCR技术分析, 发现授粉后10~24 d两糯玉米o2-NILs中6个淀粉合成基因动态表达模式不同, 其中Sh1、Sh2、SSIIIa和SBEIIb差异较大。分析胚乳转录组数据, 发现两糯玉米o2-NILs中24个海藻糖和糖基水解酶编码基因和48个o2胚乳修饰基因变化不同, 以上结果表明淀粉合成关键基因前期表达量高, 后期与对照无差异, 且糖代谢基因表达变化有利于淀粉合成可能是糯2/wx1wx1o2o2淀粉含量和百粒重不受o2突变影响, 籽粒性状明显优于黄糯2/wx1wx1o2o2的重要原因, 同时多个胚乳修饰基因的差异表达可能与该结果直接相关。本研究结果可为o2突变体在玉米育种中的应用提供重要参考。
时期
Stage | 自交系
Inbred lines | 基因型
Genotype | 赖氨酸含量
Lysine content
(%) | 可溶性糖含量
Soluble sugar
content (%) | 总淀粉含量
Starch content
(%) | 百粒重
100-kernel weight (g) | | 鲜食期 | 黄糯2 | O2O2 | 0.35 | 7.70 | 69.60 | 11.98 | | Fresh stage | Huangnuo 2 | o2o2 | 0.54** | 7.35 | 65.34** | 10.86* | | 糯2 | O2O2 | 0.42 | 6.76 | 64.11 | 16.06 | | Nuo 2 | o2o2 | 0.58** | 6.77 | 68.02** | 15.94 | | 成熟期 | 黄糯2 | O2O2 | 0.24 | 4.56 | 68.90 | 15.80 | | Mature stage | Huangnuo 2 | o2o2 | 0.43** | 5.13* | 64.60** | 14.41** | | 糯2 | O2O2 | 0.24 | 4.81 | 69.20 | 26.01 | | Nuo 2 | o2o2 | 0.38** | 5.09 | 69.10 | 25.02 |
View table in article
表1
鲜食期和成熟期糯玉米o2-NILs籽粒成分测定结果
正文中引用本图/表的段落
糯玉米是主要鲜食玉米类型, opaque2 (o2)基因导入可增加籽粒赖氨酸含量, 但同时引起籽粒皱缩、淀粉含量下降等, 限制了其育种应用。为发掘优良糯玉米受体, 以籽粒饱满圆型o2近等基因系(o2-NIL)糯2/wx1wx1o2o2和皱缩型黄糯2/wx1wx1o2o2为研究材料, 通过对鲜食期、成熟期的百粒重和籽粒成分测定, 发现淀粉和可溶性糖含量不同可能是导致2份糯玉米o2-NILs表型差异的主要原因。利用实时荧光定量PCR技术分析, 发现授粉后10~24 d两糯玉米o2-NILs中6个淀粉合成基因动态表达模式不同, 其中Sh1、Sh2、SSIIIa和SBEIIb差异较大。分析胚乳转录组数据, 发现两糯玉米o2-NILs中24个海藻糖和糖基水解酶编码基因和48个o2胚乳修饰基因变化不同, 以上结果表明淀粉合成关键基因前期表达量高, 后期与对照无差异, 且糖代谢基因表达变化有利于淀粉合成可能是糯2/wx1wx1o2o2淀粉含量和百粒重不受o2突变影响, 籽粒性状明显优于黄糯2/wx1wx1o2o2的重要原因, 同时多个胚乳修饰基因的差异表达可能与该结果直接相关。本研究结果可为o2突变体在玉米育种中的应用提供重要参考。
以Ubiquitin为内参基因, 对淀粉合成及糖代谢差异表达基因进行定量分析, 根据基因ID, 在NCBI数据库中下载基因序列。将基因序列导入Primer 5.0设计定量引物, 委托北京华大基因科技有限公司合成。将cDNA稀释作为模板, 通过普通PCR扩增目的条带, 得到的PCR产物进行琼脂糖凝胶电泳, 在紫外线照胶仪中观察条带是否特异, 本文所用定量引物信息见附表1。
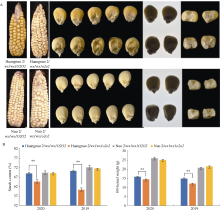
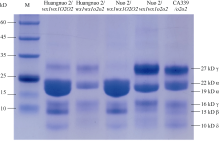
前期我们以优质蛋白玉米CA339为o2基因供体, 通过回交转育创制出多份糯玉米o2-NILs[16,25]。通过比较发现黄糯2/wx1wx1o2o2与糯2/wx1wx1o2o2籽粒表型显著不同, 黄糯2/wx1wx1o2o2为典型的o2突变体表型, 籽粒明显皱缩, 而糯2/wx1wx1o2o2籽粒呈饱满圆润状, 皱缩不明显, 但两者胚乳均呈粉质态(图1-A)。与各自轮回亲本相比(对照), 黄糯2/wx1wx1o2o2百粒重和淀粉含量显著降低, 糯2/wx1wx1o2o2籽粒则无差异, 明显高于黄糯2/wx1wx1o2o2, 并且该差异不受环境影响(图1-B)。进一步测定籽粒成分, 鲜食期与对照相比黄糯2/wx1wx1o2o2总淀粉含量和百粒重降低6.12%和1.11%, 可溶性糖含量不变, 赖氨酸含量增加54.28%; 而糯2/wx1wx1o2o2籽粒总淀粉含量增加6.12%, 赖氨酸增加38.10%, 百粒重和可溶性糖含量不受影响。这与我们前期发现的o2突变对多份糯玉米鲜食期品质影响较大, 黄糯2/wx1wx1o2o2和糯2/ wx1wx1o2o2为代表性差异材料的结果一致[27]。成熟期黄糯2/o2o2籽粒中可溶性糖增加12.50%、总淀粉含量降低6.24%, 百粒重降低8.80%, 赖氨酸含量增加79.17%; 但糯2/wx1wx1o2o2籽粒总淀粉含量、可溶性糖及百粒重均不受影响, 赖氨酸含量增加58.33% (表1)。糯2/wx1wx1o2o2籽粒表型与对照相似, 但粉质胚乳占比明显增加, 因此胚乳质地不同于QPM。黄糯2/wx1wx1o2o2胚乳中醇溶蛋白亚基27 kD γ-zein显著降低, 糯2/wx1wx1o2o2胚乳中27 kD γ-zein含量显著高于对照, 与CA339接近(图2)。此外, 15 kD β-zein、19 kD和22 kD α-zein含量在糯玉米o2-NILs均明显降低, 16 kD γ-zein在黄糯2/o2o2和糯2/wx1wx1o2o2变化相反。前期我们发现赵OP-6/wx1wx1o2o2、SY1-2/wx1wx1o2o2与糯2/wx1wx1o2o2相似, 27 kD γ-zein明显高于对照, 赵OP-6/wx1wx1o2o2淀粉含量增加, 籽粒较饱满、圆润, 而SY1-2/wx1wx1o2o2淀粉含量降低, 籽粒呈明显皱缩状[27], 表明27 kD γ-zein蛋白增加是糯玉米o2-NILs籽粒恢复饱满状的必要非充分条件, 淀粉含量也是影响籽粒饱满度的重要因素。
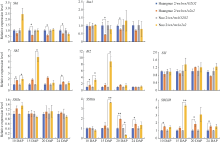
淀粉合成积累是一个持续过程, 显著影响籽粒饱满度。取授粉后10 d (10 DAP)、15 DAP、20 DAP、24 DAP胚乳对淀粉合成基因表达水平进行比较(图3)。与各自对照相比, 10 DAP时两糯玉米o2-NILs中Sh1、Sus1、SSI、SSIIa和SSIIIa表达量均无显著变化; Sh2和Bt2在 黄糯2/wx1wx1o2o2中上调1.70倍和1.55倍, 在糯2/wx1wx1o2o2中上调1.99倍和3.22倍; SBEIIb在黄糯2/wx1wx1o2o2中上调1.52倍, 在糯2/wx1wx1o2o2中无显著变化。15 DAP时, SSI和SSIIa在两糯玉米o2-NILs中无显著变化; Sh1和Sus1在黄糯2/wx1wx1o2o2中下调1.69倍和2.22倍, 但在糯2/wx1wx1o2o2中无显著变化; Sh2、Bt2、SSIIIa和SBEIIb在黄糯2/wx1wx1o2o2中无显著变化, 在糯2/wx1wx1o2o2中则分别是对照的5.79、8.88、3.63和4.27倍。20 DAP时, 与对照相比黄糯2/wx1wx1o2o2中Sh2、Bt2、SSI和SSIIa无变化, Sh1和Sus1下调1.72倍和1.30倍, SSIIIa、SBEIIb上调1.95倍和1.85倍; 糯2/wx1wx1o2o2中仅SSIIIa表达量下调3.13倍, 剩余基因均无变化。24 DAP时, 相较于对照黄糯2/wx1wx1o2o2中Sh1表达量下调1.37倍, Sh2、SSIIIa和SBEIIb上调1.48、1.45和1.54倍, Sus1、Bt2、SSI和SSIIa表达量无显著变化; 而糯2/wx1wx1o2o2中所有供试基因无变化。综上表明两糯玉米o2-NILs在授粉后15~24 d淀粉合成基因表达差异显著, 其中Sh1、Sh2、SSIIIa和SBEIIb在3个时期发生显著变化, 差异最明显, 推测是导致两糯玉米o2-NILs籽粒淀粉含量不同的主要原因。
糖代谢相关DEGs以海藻糖合成和糖基水解酶基因为主, 暗示糖代谢在两糯玉米o2-NILs中发生显著变化。动态表达分析黄糯2/wx1wx1o2o2中TPS编码基因Zm00001d032311在10 DAP时表达量无变化, 但此后上调表达, 分别上调1.37、2.71和1.75倍, 糯2/wx1wx1o2o2中10 DAP时其表达量下调1.32倍, 15 DAP上调3.38倍, 20 DAP和24 DAP时无差异。TPP编码基因Zm00001d017502在黄糯2/wx1wx1o2o2中10 DAP时表达量无变化, 15 DAP时上调4.46倍, 24 DAP时上调1.85倍, 在糯2/wx1wx1o2o2中10 DAP时表达量下调2.56倍, 20 DAP和24 DAP上调2.04倍和2.48倍; 另一TPP编码基因Zm00001d029371在黄糯2/wx1wx1o2o2中显著下调(1.40~3.33倍), 在糯2/wx1wx1o2o2中10 DAP和15 DAP下调2.86倍和3.85倍, 后2个时期不受影响。Zm00001d028243编码葡聚糖-内-1,3-β葡糖苷酶, 黄糯2/wx1wx1o2o2中15 DAP和24 DAP时表达量下调2.63和1.64倍, 但在10 DAP和20 DAP不受影响, 在糯2/wx1wx1o2o2中4个时期均下调表达(2.04~3.03倍); Zm00001d033649编码另一个糖基水解酶, 黄糯2/wx1wx1o2o2中15 DAP时表达量下调2.78倍, 其余3个时期表达量与对照无差异, 在糯2/wx1wx1o2o2中4个时期表达量均显著下调(2.50~3.45倍)。
黄糯2/wx1wx1o2o2中30个醇溶蛋白相关基因下调2.31~99.30倍(log2 FC = -1.21~ -6.63), 糯2/wx1wx1o2o2中有24个醇溶蛋白基因下调2.30~ 45.21倍(log2 FC = -1.20~ -5.50) (附表2)。其中22个为两糯玉米o2-NILs共有DEGs, 19个基因编码19 kD α-zein和22 kD α-zein, Zm00001d035760编码15 kD β-zein, 20个基因在两糯玉米o2-NILs中均下调表达, 与o2-NILs籽粒19 kD、22 kD和15 kD α-zein显著降低结果一致; Zm00001d005793编码16 kD γ-zein, 仅在黄糯2/wx1wx1o2o2中表达量下降, 与其16 kD α-zein降低一致(图2)。Zm00001d008432、Zm00001d020984、Zm00001d052079和Zm00001d 003983为赖氨酸降解基因(附表3), 其下调1.2~17.1倍(log2 FC = -1.01 ~ -4.10)使赖氨酸降解受阻, 可能是糯玉米o2-NILs赖氨酸含量增加的主要原因[28] (表1)。
玉米o2突变后籽粒表现不同, 例如o2导入自交系B46籽粒变化显著, 而导入M14籽粒基本无变化[18]。谭华等[29]将o2导入多个普通玉米自交系后发现o2-NILs赖氨酸含量大幅提高, 容重增加, 热带和亚热带种质百粒重多数下降, 但在温带种质中增加。籽粒皱缩、胚乳粉质且不透光是o2突变体的典型表现。淀粉是籽粒主要贮藏物质, 占胚乳干重的70%, 淀粉积累不足被认为是籽粒皱缩、百粒重低的重要原因。淀粉的合成是一个复杂连续过程, 授粉后10 d籽粒中淀粉开始大量合成, 20 d左右达到最大速率[30?-32], 这一过程受蔗糖合成酶(SuSy)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、淀粉合成酶(SSs)、淀粉分支酶(SBE)和去分支酶(DBE)等关键酶调控, 编码基因表达量的改变对籽粒淀粉含量影响显著[33?-35]。本研究发现黄糯2/wx1wx1o2o2和糯2/wx1wx1o2o2籽粒皱缩程度与胚乳淀粉合成基因表达不同密切相关(图1、图4和表1)。SuSy是双向催化酶, 主要功能是分解蔗糖, 为淀粉和蛋白质合成提供底物[36]。玉米K0326Y/o2o2中sh1突变, SuSy活性降低, 淀粉合成受抑制, 籽粒皱缩[37]。黄糯2/wx1wx1o2o2中Sh1表达水平显著下降, 籽粒明显皱缩, 而糯2/wx1wx1o2o2中Sh1表达不受影响, 籽粒饱满程度与对照接近。Sus1是SuSy的另一个编码基因, 表达量变化与Sh1相似, 黄糯2/wx1wx1o2o2中下调而糯2/wx1wx1o2o2中无变化, 表明黄糯2/wx1wx1o2o2胚乳中淀粉合成底物供应可能受到抑制, 但糯2/wx1wx1o2o2中不受影响。AGPase是淀粉合成限速酶, 影响碳向淀粉途径的分配[38], 胚乳中主要编码基因是Sh2和Bt2。正常籽粒授粉后蔗糖比重逐渐降低, 淀粉比重增加, 收获时以淀粉为主, 而sh2突变导致籽粒中多聚糖链合成受抑制, 蔗糖含量增加, 淀粉积累速率受抑制, 籽粒变为皱缩状[39]。10 DAP时糯2/wx1wx1o2o2和黄糯2/wx1wx1o2o2中Sh2和Bt2显著上调, 但15 DAP时仅糯2/wx1wx1o2o2中上调表达(图3), 表明淀粉积累初期(10 DAP)糯玉米o2-NILs中AGPase表达增加, 但只有糯2/wx1wx1o2o2保持较高水平至15 DAP。SSs通过催化ADPG葡萄糖基转移到葡聚糖的非还原末端来延长支链淀粉的长度, 糯玉米由于GBSS (wx)基因突变, 直链淀粉合成受抑制, 只能合成支链淀粉; SBE将α-1,4糖苷键切开, 将截短的葡聚糖链与C6羟基链接, 形成支链淀粉的分支[40]。与对照比, 两糯玉米o2-NIL中SSI、SSIIa表达水平无差异, 推测胚乳中短葡聚糖链的合成不受影响; 糯2/wx1wx1o2o2中SSIIIa和SBEIIb 15 DAP时上调表达, 与Sh2和Bt2变化一致, 表明前期糯2/wx1wx1o2o2可能积累更多支链淀粉, 与鲜食期籽粒淀粉含量高于对照相符; 黄糯2/wx1wx1o2o2中SSIIIa和SBEIIb也上调表达, 推测胚乳中淀粉合成底物不足为主要限制因素, 因此籽粒淀粉含量低于对照。根据淀粉合成基因表达模式推测, 前期(0~15 DAP)糯2/wx1wx1o2o2籽粒淀粉合成速率较高, 20 DAP时恢复至对照水平, 但黄糯2/wx1wx1o2o2自15 DAP淀粉合成持续低于对照水平。
本文的其它图/表
|