o2突变引起糯玉米籽粒淀粉积累差异研究
韩洁楠, 张泽, 刘晓丽, 李冉, 上官小川, 周婷芳, 潘越, 郝转芳, 翁建峰, 雍洪军, 周志强, 徐晶宇, 李新海, 李明顺
作物学报
2024, 50 ( 5):
1207-1222.
DOI: 10.3724/SP.J.1006.2024.33046
糯玉米是主要鲜食玉米类型, opaque2 (o2)基因导入可增加籽粒赖氨酸含量, 但同时引起籽粒皱缩、淀粉含量下降等, 限制了其育种应用。为发掘优良糯玉米受体, 以籽粒饱满圆型o2近等基因系(o2-NIL)糯2/wx1wx1o2o2和皱缩型黄糯2/wx1wx1o2o2为研究材料, 通过对鲜食期、成熟期的百粒重和籽粒成分测定, 发现淀粉和可溶性糖含量不同可能是导致2份糯玉米o2-NILs表型差异的主要原因。利用实时荧光定量PCR技术分析, 发现授粉后10~24 d两糯玉米o2-NILs中6个淀粉合成基因动态表达模式不同, 其中Sh1、Sh2、SSIIIa和SBEIIb差异较大。分析胚乳转录组数据, 发现两糯玉米o2-NILs中24个海藻糖和糖基水解酶编码基因和48个o2胚乳修饰基因变化不同, 以上结果表明淀粉合成关键基因前期表达量高, 后期与对照无差异, 且糖代谢基因表达变化有利于淀粉合成可能是糯2/wx1wx1o2o2淀粉含量和百粒重不受o2突变影响, 籽粒性状明显优于黄糯2/wx1wx1o2o2的重要原因, 同时多个胚乳修饰基因的差异表达可能与该结果直接相关。本研究结果可为o2突变体在玉米育种中的应用提供重要参考。
基因名称
Gene ID | 糯2/wx1wx1o2o2
Nuo 2/wx1wx1o2o2
log2 FC | 黄糯2/wx1wx1o2o2
Huangnuo 2/wx1wx1o2o2
log2 FC | 注释
Annotation | | Zm00001d005793 | ns | -1.21 | Prolamin 16 kD gamma zein precursor | | Zm00001d048847 | -1.80 | -5.05 | Prolamin 19 kD alpha zein z1A1_2 precursor | | Zm00001d048848 | ns | -3.56 | Prolamin 19 kD alpha zein z1A1_3 precursor | | Zm00001d048849 | ns | -3.63 | Prolamin 19 kD alpha zein z1A1_4 precursor | | Zm00001d048850 | -1.34 | -3.67 | Prolamin 19 kD alpha zein z1A1_5 precursor | | Zm00001d048851 | ns | -3.90 | Prolamin 19 kD alpha zein z1A1_6 precursor | | Zm00001d048852 | ns | -3.76 | Prolamin 19 kD alpha zein z1A1_7 precursor | | Zm00001d019155 | -1.36 | -3.70 | Prolamin 19 kD alpha zein z1B_4 precursor | | Zm00001d030855 | -1.33 | -2.18 | Prolamin 19 kD alpha zein z1D_4 precursor | | newGene_32946 | -1.67 | -2.30 | Prolamin 19 kD alpha zein z1D_2 precursor | | newGene_32956 | -2.52 | -2.73 | Prolamin 19 kD alpha zein z1D_2 precursor | | newGene_17461 | ns | -3.84 | Prolamin 19 kD alpha zein z1A2_2 precursor | | newGene_33790 | -2.19 | -3.66 | Prolamin 19 kD alpha zein z1B_1 precursor | | Zm00001d048816 | -3.78 | -4.24 | Prolamin 22 kD alpha zein z1C1_10 precursor | | Zm00001d048806 | -2.78 | -6.63 | Prolamin 22 kD alpha zein z1C1_12 precursor | | Zm00001d048817 | -5.20 | -5.80 | Prolamin 22 kD alpha zein z1C1_19 precursor | | Zm00001d048812 | -5.50 | -5.71 | Prolamin 22 kD alpha zein z1C1_7 precursor | | Zm00001d048813 | -2.75 | -3.58 | Prolamin 22 kD alpha zein z1C1_8 precursor | | Zm00001d049243 | -4.90 | -5.63 | Prolamin 22 kD alpha zein z1C2 precursor | | Zm00001d048810 | -3.69 | -4.77 | Prolamin 22 kD alpha-zein 14 | | Zm00001d048809 | -4.49 | -4.61 | Prolamin 22 kD alpha-zein 4 | | Zm00001d048818 | -4.61 | -4.30 | Prolamin 22 kD alpha-zein 8 | | Zm00001d020591 | ns | -1.87 | Prolamin 50 kD gamma zein | | Zm00001d035760 | -1.70 | ns | Prolamin PPROL 17 precursor | | Zm00001d049476 | -1.36 | -2.90 | Z1A alpha zein protein | | Zm00001d045937 | ns | -3.18 | Zein protein | | Zm00001d048807 | -4.37 | -4.79 | Zein seed storage protein, hypothetical protein | | Zm00001d019160 | -2.11 | -4.64 | Zein seed storage protein, hypothetical protein | | Zm00001d019162 | -1.49 | -5.11 | Zein seed storage protein, hypothetical protein | | Zm00001d019156 | -1.22 | -3.64 | Zein seed storage protein, hypothetical protein | | Zm00001d013100 | -1.20 | ns | Zein-binding | | Zm00001d048808 | -4.10 | -4.96 | Kafirin PSKR2 Precursor |
View table in article
附表2
糯玉米o2-NILs醇溶蛋白相关DEGs
正文中引用本图/表的段落
糯玉米是主要鲜食玉米类型, opaque2 (o2)基因导入可增加籽粒赖氨酸含量, 但同时引起籽粒皱缩、淀粉含量下降等, 限制了其育种应用。为发掘优良糯玉米受体, 以籽粒饱满圆型o2近等基因系(o2-NIL)糯2/wx1wx1o2o2和皱缩型黄糯2/wx1wx1o2o2为研究材料, 通过对鲜食期、成熟期的百粒重和籽粒成分测定, 发现淀粉和可溶性糖含量不同可能是导致2份糯玉米o2-NILs表型差异的主要原因。利用实时荧光定量PCR技术分析, 发现授粉后10~24 d两糯玉米o2-NILs中6个淀粉合成基因动态表达模式不同, 其中Sh1、Sh2、SSIIIa和SBEIIb差异较大。分析胚乳转录组数据, 发现两糯玉米o2-NILs中24个海藻糖和糖基水解酶编码基因和48个o2胚乳修饰基因变化不同, 以上结果表明淀粉合成关键基因前期表达量高, 后期与对照无差异, 且糖代谢基因表达变化有利于淀粉合成可能是糯2/wx1wx1o2o2淀粉含量和百粒重不受o2突变影响, 籽粒性状明显优于黄糯2/wx1wx1o2o2的重要原因, 同时多个胚乳修饰基因的差异表达可能与该结果直接相关。本研究结果可为o2突变体在玉米育种中的应用提供重要参考。
对多个o2突变体进行转录组分析发现, 一些基因仅在特定o2突变体/背景中表达水平有差异; 不同o2突变体中一致存在的差异表达基因, 表达水平因背景差异而明显不同[18]。玉米种内变异丰富, 因此o2突变引起的胚乳基因转录水平表达差异较大[19???-23]。Jia等[20]发现W64A/o2o2中多个淀粉合成基因转录及蛋白水平上调表达, 认为SSIIa、SBEI上调表达引起淀粉结晶度改变; Zhang等[22]发现W64A/o2o2中淀粉合成基因下调表达, 蛋白表达减少; 本课题组发现CAL58/o2o2中Sh1下调表达, Sh2、Bt2上调表达, SuSy酶活性降低, AGPase酶活性增加(数据未发表)。通过分子辅助育种可聚合o2和wx基因, 提高籽粒赖氨酸含量, 但双突自交系籽粒容重低、易感病、产量下降, 不适宜直接选育品种[24-25]。o2突变体胚乳修饰基因数量多, 多数分子机制不清楚, 缺乏相应的分子标记, 因此将o2及其修饰基因同时导入糯玉米自交系困难较大。通过比较多份糯玉米o2近等基因系(o2-NILs), 我们发现籽粒表型明显不同[16,26], 表明存在优良糯玉米o2受体, 在导入o2基因后籽粒产量与相应普通糯玉米产量接近。本研究以籽粒表型显著不同的o2-NILs黄糯2/wx1wx1o2o2和糯2/ wx1wx1o2o2为材料, 通过比较鲜食期和成熟期籽粒组成、淀粉合成基因表达和转录组差异基因等, 为发掘优良o2糯玉米受体亲本提供依据。
采用cDNA模板2 μL, 10 μL 2×SuperReal PreMix Plus, 0.6 μL引物, 6.8 μL RNase-free ddH2O配制qRT-PCR反应体系。每份样品均设置3个生物学重复和3个技术重复。使用BIO-IQ5荧光定量PCR仪进行三步法PCR反应, 程序如下: 预变性95℃ 15 min, 变性95℃ 10 s, 退火60℃ 10 s, 延伸72℃ 10 s, 40个循环。数据处理采用相对定量的分析方法采用2-ΔΔCT法, 计算公式为: 表达量比值=2-{[待测组目的基因平均CT值-待测组内参基因平均CT值]-[对照组目的基因平均CT值-对照组内参基因平均CT值]}
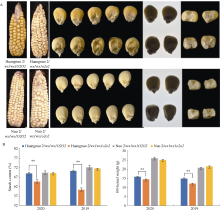
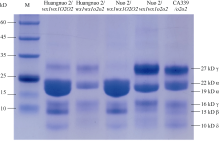
前期我们以优质蛋白玉米CA339为o2基因供体, 通过回交转育创制出多份糯玉米o2-NILs[16,25]。通过比较发现黄糯2/wx1wx1o2o2与糯2/wx1wx1o2o2籽粒表型显著不同, 黄糯2/wx1wx1o2o2为典型的o2突变体表型, 籽粒明显皱缩, 而糯2/wx1wx1o2o2籽粒呈饱满圆润状, 皱缩不明显, 但两者胚乳均呈粉质态(图1-A)。与各自轮回亲本相比(对照), 黄糯2/wx1wx1o2o2百粒重和淀粉含量显著降低, 糯2/wx1wx1o2o2籽粒则无差异, 明显高于黄糯2/wx1wx1o2o2, 并且该差异不受环境影响(图1-B)。进一步测定籽粒成分, 鲜食期与对照相比黄糯2/wx1wx1o2o2总淀粉含量和百粒重降低6.12%和1.11%, 可溶性糖含量不变, 赖氨酸含量增加54.28%; 而糯2/wx1wx1o2o2籽粒总淀粉含量增加6.12%, 赖氨酸增加38.10%, 百粒重和可溶性糖含量不受影响。这与我们前期发现的o2突变对多份糯玉米鲜食期品质影响较大, 黄糯2/wx1wx1o2o2和糯2/ wx1wx1o2o2为代表性差异材料的结果一致[27]。成熟期黄糯2/o2o2籽粒中可溶性糖增加12.50%、总淀粉含量降低6.24%, 百粒重降低8.80%, 赖氨酸含量增加79.17%; 但糯2/wx1wx1o2o2籽粒总淀粉含量、可溶性糖及百粒重均不受影响, 赖氨酸含量增加58.33% (表1)。糯2/wx1wx1o2o2籽粒表型与对照相似, 但粉质胚乳占比明显增加, 因此胚乳质地不同于QPM。黄糯2/wx1wx1o2o2胚乳中醇溶蛋白亚基27 kD γ-zein显著降低, 糯2/wx1wx1o2o2胚乳中27 kD γ-zein含量显著高于对照, 与CA339接近(图2)。此外, 15 kD β-zein、19 kD和22 kD α-zein含量在糯玉米o2-NILs均明显降低, 16 kD γ-zein在黄糯2/o2o2和糯2/wx1wx1o2o2变化相反。前期我们发现赵OP-6/wx1wx1o2o2、SY1-2/wx1wx1o2o2与糯2/wx1wx1o2o2相似, 27 kD γ-zein明显高于对照, 赵OP-6/wx1wx1o2o2淀粉含量增加, 籽粒较饱满、圆润, 而SY1-2/wx1wx1o2o2淀粉含量降低, 籽粒呈明显皱缩状[27], 表明27 kD γ-zein蛋白增加是糯玉米o2-NILs籽粒恢复饱满状的必要非充分条件, 淀粉含量也是影响籽粒饱满度的重要因素。
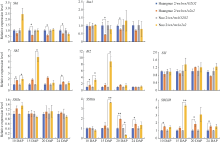
淀粉合成积累是一个持续过程, 显著影响籽粒饱满度。取授粉后10 d (10 DAP)、15 DAP、20 DAP、24 DAP胚乳对淀粉合成基因表达水平进行比较(图3)。与各自对照相比, 10 DAP时两糯玉米o2-NILs中Sh1、Sus1、SSI、SSIIa和SSIIIa表达量均无显著变化; Sh2和Bt2在 黄糯2/wx1wx1o2o2中上调1.70倍和1.55倍, 在糯2/wx1wx1o2o2中上调1.99倍和3.22倍; SBEIIb在黄糯2/wx1wx1o2o2中上调1.52倍, 在糯2/wx1wx1o2o2中无显著变化。15 DAP时, SSI和SSIIa在两糯玉米o2-NILs中无显著变化; Sh1和Sus1在黄糯2/wx1wx1o2o2中下调1.69倍和2.22倍, 但在糯2/wx1wx1o2o2中无显著变化; Sh2、Bt2、SSIIIa和SBEIIb在黄糯2/wx1wx1o2o2中无显著变化, 在糯2/wx1wx1o2o2中则分别是对照的5.79、8.88、3.63和4.27倍。20 DAP时, 与对照相比黄糯2/wx1wx1o2o2中Sh2、Bt2、SSI和SSIIa无变化, Sh1和Sus1下调1.72倍和1.30倍, SSIIIa、SBEIIb上调1.95倍和1.85倍; 糯2/wx1wx1o2o2中仅SSIIIa表达量下调3.13倍, 剩余基因均无变化。24 DAP时, 相较于对照黄糯2/wx1wx1o2o2中Sh1表达量下调1.37倍, Sh2、SSIIIa和SBEIIb上调1.48、1.45和1.54倍, Sus1、Bt2、SSI和SSIIa表达量无显著变化; 而糯2/wx1wx1o2o2中所有供试基因无变化。综上表明两糯玉米o2-NILs在授粉后15~24 d淀粉合成基因表达差异显著, 其中Sh1、Sh2、SSIIIa和SBEIIb在3个时期发生显著变化, 差异最明显, 推测是导致两糯玉米o2-NILs籽粒淀粉含量不同的主要原因。
为进一步探究o2突变导致黄糯2/wx1wx1o2o2和糯2/wx1wx1o2o2籽粒淀粉积累不同原因, 对两材料20 DAP胚乳进行转录组测序。以各自轮回亲本为对照分析黄糯2/wx1wx1o2o2和糯2/wx1wx1o2o2差异表达基因(differently expressed genes, DEGs), 黄糯2/wx1wx1o2o2中共有2466个DEGs, 其中1290个基因上调表达, 1176个基因下调表达; 糯2/wx1wx1o2o2中共有1685个DEGs, 其中1124个基因上调表达, 561个基因下调表达(图4-A)。糯2/wx1wx1o2o2中DEGs远低于黄糯2/wx1wx1o2o2, 表明o2突变对胚乳发育的影响依赖于基因型, 糯2/wx1wx1o2o2受影响程度可能低于黄糯2/wx1wx1o2o2。对DEGs进行KEGG路径富集, 黄糯2/wx1wx1o2o2 DEGs主要富集到内质网蛋白质加工、碳代谢、DNA复制、淀粉和蔗糖代谢路径(图4-B); 糯2/wx1wx1o2o2主要富集到碳代谢、淀粉和蔗糖代谢、甘氨酸、丝氨酸和苏氨酸代谢、苯丙烷生物合成和糖酵解5个路径(图4-C)。
(A): 两糯玉米o2-NILs差异表达基因数目; (B): 黄糯2/wx1wx1o2o2差异表达基因KEGG富集分析; (C): 糯2/wx1wx1o2o2差异表达基因KEGG富集分析。
授粉后20 d两糯玉米o2-NILs共筛选出34个淀粉和糖代谢相关DEGs (图5-A), 淀粉合成相关DEGs有AGPase小亚基、淀粉合成酶SSIII、葡萄糖变位酶、异淀粉酶和α淀粉酶编码基因共计7个。剩余27个DEGs为糖代谢相关基因(图5-B), 其中4个DEGs为两糯玉米o2-NILs共有基因, 分别编码丙酮酸磷酸双激酶(PPDK)、海藻糖磷酸合酶(TPP)和果胶酯酶, 在两糯玉米o2-NILs中一致下调。15个DEGs仅在黄糯2/wx1wx1o2o2中差异表达, 其中12个基因下调, 3个基因上调, 包括5个海藻糖六磷酸合酶(TPS)基因、3个TPP基因、4个糖基水解酶基因、2个果糖激酶、1个果胶酯酶; 7个DEGs在糯2/wx1wx1o2o2中特异表达, 其中5个基因下调表达, 2个基因上调, 包括2个TPP编码基因和5个糖基水解酶基因。对8个DEGs进行RT-PCR验证, 发现与RNA-seq结果一致(图6)。
糖代谢相关DEGs以海藻糖合成和糖基水解酶基因为主, 暗示糖代谢在两糯玉米o2-NILs中发生显著变化。动态表达分析黄糯2/wx1wx1o2o2中TPS编码基因Zm00001d032311在10 DAP时表达量无变化, 但此后上调表达, 分别上调1.37、2.71和1.75倍, 糯2/wx1wx1o2o2中10 DAP时其表达量下调1.32倍, 15 DAP上调3.38倍, 20 DAP和24 DAP时无差异。TPP编码基因Zm00001d017502在黄糯2/wx1wx1o2o2中10 DAP时表达量无变化, 15 DAP时上调4.46倍, 24 DAP时上调1.85倍, 在糯2/wx1wx1o2o2中10 DAP时表达量下调2.56倍, 20 DAP和24 DAP上调2.04倍和2.48倍; 另一TPP编码基因Zm00001d029371在黄糯2/wx1wx1o2o2中显著下调(1.40~3.33倍), 在糯2/wx1wx1o2o2中10 DAP和15 DAP下调2.86倍和3.85倍, 后2个时期不受影响。Zm00001d028243编码葡聚糖-内-1,3-β葡糖苷酶, 黄糯2/wx1wx1o2o2中15 DAP和24 DAP时表达量下调2.63和1.64倍, 但在10 DAP和20 DAP不受影响, 在糯2/wx1wx1o2o2中4个时期均下调表达(2.04~3.03倍); Zm00001d033649编码另一个糖基水解酶, 黄糯2/wx1wx1o2o2中15 DAP时表达量下调2.78倍, 其余3个时期表达量与对照无差异, 在糯2/wx1wx1o2o2中4个时期表达量均显著下调(2.50~3.45倍)。
黄糯2/wx1wx1o2o2中30个醇溶蛋白相关基因下调2.31~99.30倍(log2 FC = -1.21~ -6.63), 糯2/wx1wx1o2o2中有24个醇溶蛋白基因下调2.30~ 45.21倍(log2 FC = -1.20~ -5.50) (附表2)。其中22个为两糯玉米o2-NILs共有DEGs, 19个基因编码19 kD α-zein和22 kD α-zein, Zm00001d035760编码15 kD β-zein, 20个基因在两糯玉米o2-NILs中均下调表达, 与o2-NILs籽粒19 kD、22 kD和15 kD α-zein显著降低结果一致; Zm00001d005793编码16 kD γ-zein, 仅在黄糯2/wx1wx1o2o2中表达量下降, 与其16 kD α-zein降低一致(图2)。Zm00001d008432、Zm00001d020984、Zm00001d052079和Zm00001d 003983为赖氨酸降解基因(附表3), 其下调1.2~17.1倍(log2 FC = -1.01 ~ -4.10)使赖氨酸降解受阻, 可能是糯玉米o2-NILs赖氨酸含量增加的主要原因[28] (表1)。
玉米o2突变后籽粒表现不同, 例如o2导入自交系B46籽粒变化显著, 而导入M14籽粒基本无变化[18]。谭华等[29]将o2导入多个普通玉米自交系后发现o2-NILs赖氨酸含量大幅提高, 容重增加, 热带和亚热带种质百粒重多数下降, 但在温带种质中增加。籽粒皱缩、胚乳粉质且不透光是o2突变体的典型表现。淀粉是籽粒主要贮藏物质, 占胚乳干重的70%, 淀粉积累不足被认为是籽粒皱缩、百粒重低的重要原因。淀粉的合成是一个复杂连续过程, 授粉后10 d籽粒中淀粉开始大量合成, 20 d左右达到最大速率[30?-32], 这一过程受蔗糖合成酶(SuSy)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、淀粉合成酶(SSs)、淀粉分支酶(SBE)和去分支酶(DBE)等关键酶调控, 编码基因表达量的改变对籽粒淀粉含量影响显著[33?-35]。本研究发现黄糯2/wx1wx1o2o2和糯2/wx1wx1o2o2籽粒皱缩程度与胚乳淀粉合成基因表达不同密切相关(图1、图4和表1)。SuSy是双向催化酶, 主要功能是分解蔗糖, 为淀粉和蛋白质合成提供底物[36]。玉米K0326Y/o2o2中sh1突变, SuSy活性降低, 淀粉合成受抑制, 籽粒皱缩[37]。黄糯2/wx1wx1o2o2中Sh1表达水平显著下降, 籽粒明显皱缩, 而糯2/wx1wx1o2o2中Sh1表达不受影响, 籽粒饱满程度与对照接近。Sus1是SuSy的另一个编码基因, 表达量变化与Sh1相似, 黄糯2/wx1wx1o2o2中下调而糯2/wx1wx1o2o2中无变化, 表明黄糯2/wx1wx1o2o2胚乳中淀粉合成底物供应可能受到抑制, 但糯2/wx1wx1o2o2中不受影响。AGPase是淀粉合成限速酶, 影响碳向淀粉途径的分配[38], 胚乳中主要编码基因是Sh2和Bt2。正常籽粒授粉后蔗糖比重逐渐降低, 淀粉比重增加, 收获时以淀粉为主, 而sh2突变导致籽粒中多聚糖链合成受抑制, 蔗糖含量增加, 淀粉积累速率受抑制, 籽粒变为皱缩状[39]。10 DAP时糯2/wx1wx1o2o2和黄糯2/wx1wx1o2o2中Sh2和Bt2显著上调, 但15 DAP时仅糯2/wx1wx1o2o2中上调表达(图3), 表明淀粉积累初期(10 DAP)糯玉米o2-NILs中AGPase表达增加, 但只有糯2/wx1wx1o2o2保持较高水平至15 DAP。SSs通过催化ADPG葡萄糖基转移到葡聚糖的非还原末端来延长支链淀粉的长度, 糯玉米由于GBSS (wx)基因突变, 直链淀粉合成受抑制, 只能合成支链淀粉; SBE将α-1,4糖苷键切开, 将截短的葡聚糖链与C6羟基链接, 形成支链淀粉的分支[40]。与对照比, 两糯玉米o2-NIL中SSI、SSIIa表达水平无差异, 推测胚乳中短葡聚糖链的合成不受影响; 糯2/wx1wx1o2o2中SSIIIa和SBEIIb 15 DAP时上调表达, 与Sh2和Bt2变化一致, 表明前期糯2/wx1wx1o2o2可能积累更多支链淀粉, 与鲜食期籽粒淀粉含量高于对照相符; 黄糯2/wx1wx1o2o2中SSIIIa和SBEIIb也上调表达, 推测胚乳中淀粉合成底物不足为主要限制因素, 因此籽粒淀粉含量低于对照。根据淀粉合成基因表达模式推测, 前期(0~15 DAP)糯2/wx1wx1o2o2籽粒淀粉合成速率较高, 20 DAP时恢复至对照水平, 但黄糯2/wx1wx1o2o2自15 DAP淀粉合成持续低于对照水平。
淀粉的合成还与糖代谢密切相关, 例如海藻糖合成、糖酵解等均可与淀粉合成底物或中间产物相互转化。海藻糖-6-磷酸酶(TPS)催化UDPG和葡萄糖-6-磷酸(G6P)合成海藻糖-6-磷酸(T6P), 进而在海藻糖-6-磷酸磷酸酶(TPP)作用下合成海藻糖。G6P和UDPG是淀粉合成和碳代谢底物, T6P为G6P和UDPG库容标志物, 在植物生长及碳利用中有重要作用[41]。豌豆缺乏T6P, 种子大小和淀粉含量受影响, 表型与孟德尔研究的皱缩豌豆种子相似[42]; 过表达大肠杆菌TPS基因可增加拟南芥叶片淀粉合成[43]; Hu等[44]发现增强TPS或降低TPP活性可增加玉米籽粒淀粉含量。T6P还与蔗糖非发酵蛋白激酶(SnRK1)互作双向调控蔗糖和淀粉合成[45]。植物TPS和TPP酶由多个基因编码[46], 20 DAP时糯2/wx1wx1o2o2中TPS基因表达不受影响, 多个TPP基因下调, 推测糯2/wx1wx1o2o2 TPS活性不受影响, 但TPP活性降低, 有利于淀粉合成; 而黄糯2/wx1wx1o2o2中多数TPS和TPP编码基因下调, 推测黄糯2/wx1wx1o2o2 TPS和TPP活性均降低, 不利于淀粉生成(图5-B)。进一步分析Zm00001d032311和Zm00001d017502动态表达模式说明海藻糖途径少数基因的变化与淀粉合成速率直接相关(图6)。果糖激酶在糖酵解途径发挥重要作用, 可与蔗糖合酶协同调控蔗糖合成与降解[47], 番茄幼果中通过调节蔗糖输入, 对淀粉积累产生影响[48-49]。本研究发现o2突变仅引起2个果糖激酶基因在黄糯2中下调表达, 是两糯玉米o2-NILs糖酵解受影响不同的表现。结构多样的糖苷水解酶在糖和糖缀合物水解与合成中扮演重要角色, 编码基因众多, 例如拟南芥有393个编码基因[50]。α-葡萄糖苷酶参与淀粉及糖原代谢, β-木糖酶与β-葡萄糖苷酶为纤维素酶, 降解纤维素生成纤维二糖和葡萄糖[51], 以上编码基因在两糯玉米o2-NILs多数下调, 说明o2突变可能抑制了胚乳的糖苷水解, 使胚乳发育和细胞壁代谢受影响。果胶是大分子多糖, 与植物生长发育、逆境应答等生物学过程密切联系[52], 番茄中催化果胶多聚体形成的半乳糖醛酸转移酶编码基因GAUT4突变, 果实果胶结构改变, 淀粉和果实含量降低[53]。果胶酯酶是三大果胶酶之一, 随机切除水溶性果胶分子的酯键, 产生游离羧基[54], 苹果果胶甲酯酶MdPME2在粉质化不同的果实中呈现表达差异[55]。o2突变导致果胶酯酶基因下调表达, 可能与糯玉米o2-NILs胚乳质地改变相关。磷酸丙酮酸双激酶(PPDK)是CO2固定关键酶, 胚乳中高表达PPDK可与淀粉合成酶稳定结合, 协同调控后期籽粒灌浆[56], 两糯玉米o2-NILs中PPDK1一致下调, 可能导致淀粉酶活性降低。
醇溶蛋白形成后储存于蛋白体中, 若醇溶蛋白与淀粉同步合成, 则紧密结合形成硬质胚乳占比高的硬粒玉米。糯玉米o2-NILs醇溶蛋白合成受阻, 淀粉合成基因也受影响(附表2和图3), 导致成熟籽粒中储藏物质降低, 胚乳几乎全部变为粉质状(图1)。27-kD γ-zein调控蛋白体形成数量, QPM胚乳中γ27蛋白表达量增加, 是公认的o2修饰基因, 并且o2修饰基因数量与γ27蛋白表达正相关[57-58], 糯2/wx1wx1o2o2中27 kD γ-zein表达量高于黄糯2/wx1wx1o2o2 (图2), 暗示糯2/wx1wx1o2o2中胚乳修饰基因数量多。通过定位群体Holding发掘到多个胚乳修饰基因位点, 主效基因位于7号染色体; 包括27-kD γ-zein、葡萄糖转运蛋白、焦磷酸依赖型果糖-6-磷酸1-磷酸转移酶α亚基(PFPα)、蛋白质磷酸酶2C (PP2C)等16个基因位于连锁区间内, 这些基因在QPM中多数上调表达[59-60]。Li等[61]发现热激蛋白(HSPs)、HSP伴侣蛋白(ERDJ3A)、丝氨酸/精氨酸富含蛋白质编码基因(SR45a)在QPM中也呈上调表达。本研究发现黄糯2/wx1wx1o2o2中39个o2胚乳修饰基因差异表达, 包括32个HSPs、5个PP2C、1个ERDJ3A和1个SR45a, 其中仅4个基因上调; 糯2/wx1wx1o2o2中13个o2胚乳修饰基因差异表达, 包括6个HSPs、6个PP2C和1个PFPα, 其中8个基因上调(附表4)。PFPα表达增加使PFP催化活性提高, 加速糖酵解, 改善o2突变引起的能量匮乏, 与硬质透明胚乳恢复相关[59??-62]; SR45a参与RNA剪切, 拟南芥中受胁迫时其上调表达[63], 水稻中ERDJ3A在内质网应激下被激活, 可促进未折叠蛋白在液泡和内质网之间的传递[64]。上述基因在糯玉米o2-NILs均下调表达, 是籽粒表型不同于普通糯玉米的重要原因。PP2C功能与脱落酸信号途径有关, 在优质蛋白玉米K0326Y/o2o2中上调, 可能与其他修饰基因共同起作用[57], 但在糯玉米o2-NILs变化趋势不一致。HSPs在QPM中上调, 对缓解o2突变引起未折叠单重组或蛋白质聚集引起的应激效应有重要作用[61-62], 糯2/wx1wx1o2o2中仅1个HSP下调, 而黄糯2/wx1wx1o2o2中有30个, 这可能与糯2/wx1wx1o2o2籽粒淀粉和百粒重不受影响直接相关, 需进一步研究。
将o2基因导入糯玉米可显著提高籽粒赖氨酸含量, 改善营养价值, 但与o2基因连锁的不良性状限制了优质蛋白糯玉米选育。与皱缩型o2近等基因系黄糯2/wx1wx1o2o2相比, 糯2/wx1wx1o2o2玉米成熟期淀粉含量及百粒重明显提高, 籽粒圆润饱满。研究发现糯2/wx1wx1o2o2与黄糯2/wx1wx1o2o2中以Sh1、Sh2、SSIIIa和SBEIIb为主的淀粉合成基因、以海藻糖和糖基水解酶为主的糖代谢基因表达模式明显不同, 推测糯2/wx1wx1o2o2 10~15 DAP淀粉合成速率高, 后与对照无差异, 但黄糯2/wx1wx1o2o2自15 DAP起淀粉合成速率持续低于对照, 且糯2/wx1wx1o2o2中糖代谢基因的变化更有利于淀粉合成。根据转录组差异表达基因分析结果, 认为胚乳修饰基因, 尤其是热激蛋白编码基因的差异表达可能与两糯玉米o2-NILs籽粒淀粉和百粒重不同直接相关。
本文的其它图/表
|