o2突变引起糯玉米籽粒淀粉积累差异研究
韩洁楠, 张泽, 刘晓丽, 李冉, 上官小川, 周婷芳, 潘越, 郝转芳, 翁建峰, 雍洪军, 周志强, 徐晶宇, 李新海, 李明顺
作物学报
2024, 50 ( 5):
1207-1222.
DOI: 10.3724/SP.J.1006.2024.33046
糯玉米是主要鲜食玉米类型, opaque2 (o2)基因导入可增加籽粒赖氨酸含量, 但同时引起籽粒皱缩、淀粉含量下降等, 限制了其育种应用。为发掘优良糯玉米受体, 以籽粒饱满圆型o2近等基因系(o2-NIL)糯2/wx1wx1o2o2和皱缩型黄糯2/wx1wx1o2o2为研究材料, 通过对鲜食期、成熟期的百粒重和籽粒成分测定, 发现淀粉和可溶性糖含量不同可能是导致2份糯玉米o2-NILs表型差异的主要原因。利用实时荧光定量PCR技术分析, 发现授粉后10~24 d两糯玉米o2-NILs中6个淀粉合成基因动态表达模式不同, 其中Sh1、Sh2、SSIIIa和SBEIIb差异较大。分析胚乳转录组数据, 发现两糯玉米o2-NILs中24个海藻糖和糖基水解酶编码基因和48个o2胚乳修饰基因变化不同, 以上结果表明淀粉合成关键基因前期表达量高, 后期与对照无差异, 且糖代谢基因表达变化有利于淀粉合成可能是糯2/wx1wx1o2o2淀粉含量和百粒重不受o2突变影响, 籽粒性状明显优于黄糯2/wx1wx1o2o2的重要原因, 同时多个胚乳修饰基因的差异表达可能与该结果直接相关。本研究结果可为o2突变体在玉米育种中的应用提供重要参考。

View image in article
图2
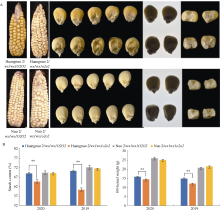
糯玉米胚乳醇溶蛋白亚基分析
正文中引用本图/表的段落
前期我们以优质蛋白玉米CA339为o2基因供体, 通过回交转育创制出多份糯玉米o2-NILs[16,25]。通过比较发现黄糯2/wx1wx1o2o2与糯2/wx1wx1o2o2籽粒表型显著不同, 黄糯2/wx1wx1o2o2为典型的o2突变体表型, 籽粒明显皱缩, 而糯2/wx1wx1o2o2籽粒呈饱满圆润状, 皱缩不明显, 但两者胚乳均呈粉质态(图1-A)。与各自轮回亲本相比(对照), 黄糯2/wx1wx1o2o2百粒重和淀粉含量显著降低, 糯2/wx1wx1o2o2籽粒则无差异, 明显高于黄糯2/wx1wx1o2o2, 并且该差异不受环境影响(图1-B)。进一步测定籽粒成分, 鲜食期与对照相比黄糯2/wx1wx1o2o2总淀粉含量和百粒重降低6.12%和1.11%, 可溶性糖含量不变, 赖氨酸含量增加54.28%; 而糯2/wx1wx1o2o2籽粒总淀粉含量增加6.12%, 赖氨酸增加38.10%, 百粒重和可溶性糖含量不受影响。这与我们前期发现的o2突变对多份糯玉米鲜食期品质影响较大, 黄糯2/wx1wx1o2o2和糯2/ wx1wx1o2o2为代表性差异材料的结果一致[27]。成熟期黄糯2/o2o2籽粒中可溶性糖增加12.50%、总淀粉含量降低6.24%, 百粒重降低8.80%, 赖氨酸含量增加79.17%; 但糯2/wx1wx1o2o2籽粒总淀粉含量、可溶性糖及百粒重均不受影响, 赖氨酸含量增加58.33% (表1)。糯2/wx1wx1o2o2籽粒表型与对照相似, 但粉质胚乳占比明显增加, 因此胚乳质地不同于QPM。黄糯2/wx1wx1o2o2胚乳中醇溶蛋白亚基27 kD γ-zein显著降低, 糯2/wx1wx1o2o2胚乳中27 kD γ-zein含量显著高于对照, 与CA339接近(图2)。此外, 15 kD β-zein、19 kD和22 kD α-zein含量在糯玉米o2-NILs均明显降低, 16 kD γ-zein在黄糯2/o2o2和糯2/wx1wx1o2o2变化相反。前期我们发现赵OP-6/wx1wx1o2o2、SY1-2/wx1wx1o2o2与糯2/wx1wx1o2o2相似, 27 kD γ-zein明显高于对照, 赵OP-6/wx1wx1o2o2淀粉含量增加, 籽粒较饱满、圆润, 而SY1-2/wx1wx1o2o2淀粉含量降低, 籽粒呈明显皱缩状[27], 表明27 kD γ-zein蛋白增加是糯玉米o2-NILs籽粒恢复饱满状的必要非充分条件, 淀粉含量也是影响籽粒饱满度的重要因素。
黄糯2/wx1wx1o2o2中30个醇溶蛋白相关基因下调2.31~99.30倍(log2 FC = -1.21~ -6.63), 糯2/wx1wx1o2o2中有24个醇溶蛋白基因下调2.30~ 45.21倍(log2 FC = -1.20~ -5.50) (附表2)。其中22个为两糯玉米o2-NILs共有DEGs, 19个基因编码19 kD α-zein和22 kD α-zein, Zm00001d035760编码15 kD β-zein, 20个基因在两糯玉米o2-NILs中均下调表达, 与o2-NILs籽粒19 kD、22 kD和15 kD α-zein显著降低结果一致; Zm00001d005793编码16 kD γ-zein, 仅在黄糯2/wx1wx1o2o2中表达量下降, 与其16 kD α-zein降低一致(图2)。Zm00001d008432、Zm00001d020984、Zm00001d052079和Zm00001d 003983为赖氨酸降解基因(附表3), 其下调1.2~17.1倍(log2 FC = -1.01 ~ -4.10)使赖氨酸降解受阻, 可能是糯玉米o2-NILs赖氨酸含量增加的主要原因[28] (表1)。
醇溶蛋白形成后储存于蛋白体中, 若醇溶蛋白与淀粉同步合成, 则紧密结合形成硬质胚乳占比高的硬粒玉米。糯玉米o2-NILs醇溶蛋白合成受阻, 淀粉合成基因也受影响(附表2和图3), 导致成熟籽粒中储藏物质降低, 胚乳几乎全部变为粉质状(图1)。27-kD γ-zein调控蛋白体形成数量, QPM胚乳中γ27蛋白表达量增加, 是公认的o2修饰基因, 并且o2修饰基因数量与γ27蛋白表达正相关[57-58], 糯2/wx1wx1o2o2中27 kD γ-zein表达量高于黄糯2/wx1wx1o2o2 (图2), 暗示糯2/wx1wx1o2o2中胚乳修饰基因数量多。通过定位群体Holding发掘到多个胚乳修饰基因位点, 主效基因位于7号染色体; 包括27-kD γ-zein、葡萄糖转运蛋白、焦磷酸依赖型果糖-6-磷酸1-磷酸转移酶α亚基(PFPα)、蛋白质磷酸酶2C (PP2C)等16个基因位于连锁区间内, 这些基因在QPM中多数上调表达[59-60]。Li等[61]发现热激蛋白(HSPs)、HSP伴侣蛋白(ERDJ3A)、丝氨酸/精氨酸富含蛋白质编码基因(SR45a)在QPM中也呈上调表达。本研究发现黄糯2/wx1wx1o2o2中39个o2胚乳修饰基因差异表达, 包括32个HSPs、5个PP2C、1个ERDJ3A和1个SR45a, 其中仅4个基因上调; 糯2/wx1wx1o2o2中13个o2胚乳修饰基因差异表达, 包括6个HSPs、6个PP2C和1个PFPα, 其中8个基因上调(附表4)。PFPα表达增加使PFP催化活性提高, 加速糖酵解, 改善o2突变引起的能量匮乏, 与硬质透明胚乳恢复相关[59??-62]; SR45a参与RNA剪切, 拟南芥中受胁迫时其上调表达[63], 水稻中ERDJ3A在内质网应激下被激活, 可促进未折叠蛋白在液泡和内质网之间的传递[64]。上述基因在糯玉米o2-NILs均下调表达, 是籽粒表型不同于普通糯玉米的重要原因。PP2C功能与脱落酸信号途径有关, 在优质蛋白玉米K0326Y/o2o2中上调, 可能与其他修饰基因共同起作用[57], 但在糯玉米o2-NILs变化趋势不一致。HSPs在QPM中上调, 对缓解o2突变引起未折叠单重组或蛋白质聚集引起的应激效应有重要作用[61-62], 糯2/wx1wx1o2o2中仅1个HSP下调, 而黄糯2/wx1wx1o2o2中有30个, 这可能与糯2/wx1wx1o2o2籽粒淀粉和百粒重不受影响直接相关, 需进一步研究。
本文的其它图/表
|




{kind=link}