Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2022, Vol. 48 ›› Issue (1): 249-258.doi: 10.3724/SP.J.1006.2022.14019
• RESEARCH NOTES • Previous Articles Next Articles
YU Hui-Fang( ), ZHANG Wei-Na, KANG Yi-Chen, FAN Yan-Ling, YANG Xin-Yu, SHI Ming-Fu, ZHANG Ru-Yan, ZHANG Jun-Lian, QIN Shu-Hao*()
), ZHANG Wei-Na, KANG Yi-Chen, FAN Yan-Ling, YANG Xin-Yu, SHI Ming-Fu, ZHANG Ru-Yan, ZHANG Jun-Lian, QIN Shu-Hao*()
| [1] | Liu H, Xue X, Yu Y, Xu M, Lu C, Meng X, Zhang B, Ding X, Chu Z. Copper ions suppress abscisic acid biosynthesis to enhance defence against phytophthora infectants in potato. Mol Plant Pathol, 2020, 21:636-651. |
| [2] | Farber D H, Koga C, Johnson D A. Disease gradients of late blight of potato from infrared images of commercial fields. Am J Potato Res, 2020, 97:347-359. |
| [3] | Gish L A, Clark S E. The RLK/Pelle family of kinases. Plant J, 2011, 66:117-127. |
| [4] | Walker J C, Zhang R. Relationship of a putative receptor protein kinase from maize to the S-locus glycoproteins of Brassica. Nature, 1990, 345:743-746. |
| [5] | Shiu S H, Karlowski W M, Pan R, Tzeng Y H, Mayer K F X, Li W H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell, 2004, 16:1220-1234. |
| [6] | Shiu S H, Bleecker A B. Receptor-like kinases from Arabidopsis form a mono-phyletic gene family related to animal receptor kinases. Proc Natl Acad Sci USA, 2001, 98:10763-10768. |
| [7] | Han Y F, Yang Q, Zhang S W, Sun D Y, Sun Y. Receptor like kinase CrRLK1-L subfamily: novel motifs in extracellular domain and biological functions in plants. Prog Biochem Biophys, 2011, 38:891-899. |
| [8] | Schulze-Muth P, Irmler S, Schröder G, Schröder J. Novel type of receptor-like protein kinase from a higher plant (Catharanthus roseus): cDNA, gene, intramolecular autophosphorylation, and identification of a threonine important for auto- and substrate phosphorylation. J Biol Chem, 1996, 271:26684-26689. |
| [9] | Lindner H, Muller L M, Boisson-Dernier A, Grossniklaus U. CrRLK1L receptor-like kinases: not just another brick in thewall. Curr Opin Plant Biol, 2012, 6:659-669. |
| [10] | Boisson-Dernier A, Kessler S A, Grossniklaus U. The walls have ears: the role of plant CrRLK1Ls in sensingand transducing extracellular signalsyigw. J Exp Bot, 2011, 62:1581-1591. |
| [11] | 韩永峰, 杨倩, 张胜伟, 孙大业, 孙颖. 植物类受体激酶 CrRLK1-L 亚家族及其生物学功能. 生物化学与生物物理进展, 2011, 38:891-899. |
| Han Y F, Yang Q, Zhang S W, Sun D Y, Sun Y. The biological functions of Plant receptor kinase CRRLK1-L subfamily. Prog Biochem Biophys, 2011, 38:891-899 (in Chinese with English abstract). | |
| [12] | Greeff C C, Roux M M, Mundy J J, Petersen M M. Receptor-like kinase complexes in plant innate immunity. Front Plant Sci, 2012, 3:209. |
| [13] | Fujita M, Fujita Y, Noutoshi Y, Takahashi F, Narusaka Y, Yamaguchi-Shinozaki K, Shinozaki K. Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks. Curr Opin Plant Biol, 2006, 9:436-442. |
| [14] | Nguyen Q N, Lee Y S, Cho L H, Jeong H J, An G, Jung K H. Genome-wide identification and analysis of Catharanthus roseus RLK1-like kinases in rice. Planta, 2015, 241:603-613. |
| [15] | Zuo C W, Zhang W N, Ma Z H, Chu M Y, Mao J, An Z S, Chen B H. Genome-wide identification and expression analysis of the CrRLK1L gene family in apple (Malus domestica). Plant Mol Biol Rep, 2018, 36:844-857. |
| [16] | Kou X B, Qi K J, Qiao X, Yin H, Liu X, Zhang S L, Wu J Y. Evolution, expression analysis, and functional verification of Catharanthus roseus RLK1-like kinase (CrRLK1L) family proteins in pear (Pyrus bretchneideri). Genomics, 2017, 109:290-301. |
| [17] | Guo H, Nolan T M, Song G, Liu S, Xie Z, Chen J, Schnable P S, Walley J W, Yin Y. FERONIA receptor kinase contributes to plant immunity by suppressing jasmonic acid signaling in Arabidopsis thaliana. Curr Biol, 2018, 28:3316-3324. |
| [18] | Kanaoka M M, Torii K U. FERONIA as an upstream receptor kinase for polar cell growth in plants. Proc Natl Acad Sci USA, 2010, 107:17461-17462. |
| [19] | Richter J, Ploderer M, Mongelard G, Gutierrez L, Hauser M T. Role of CrRLK1L cell wall sensors HERCULES1 and 2, THESEUS1, and FERONIA in growth adaptation triggered by heavy metals and trace elements. Front Plant Sci, 2017, 8:1554. |
| [20] | Miyazaki S, Murata T, Sakurai-Ozato N, Kubo M, Demura T, Fukuda H, Hasebe M. ANXUR1 and 2, sister genes to FERONIA/SIRENE, are male factors for coordinated fertilization. Curr Biol, 2009, 19:1327-1331. |
| [21] | Yang Z, Xing J, Wang L, Liu Y, Qu J, Tan Y, Fu X, Lin Q, Deng H, Yu F. Mutations of two FERONIA-like receptor genes enhance rice blast resistance without growth penalty. J Exp Bot, 2020, 71:2112-2126. |
| [22] | Liu R H, Meng J L. MapDraw: a microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Hereditas, 2003, 25:317-321. |
| [23] | Thompson J D, Gibson T J, Plewniak F, Jeanmougin F, Higgins D G. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res, 1997, 25:4876-4882. |
| [24] | 蒋锐. 马铃薯晚疫病广谱抗性QTL dPI09c的精细定位及抗性基因克隆. 华中农业大学博士学位论文,湖北武汉, 2017. |
| Jiang R. Fine Mapping, Cloning and Function Dissection of the Gene Conferring Durable Late Blight Resistance of QTL dPI09c in Potato. PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei,China, 2017 (in Chinese with English abstract). | |
| [25] | 巩檑, 甘晓燕, 张丽, 陈虞超, 聂峰杰, 石磊, 郭志乾, 宋玉霞. 马铃薯StNAC72 基因克隆及表达分析. 分子植物育种, 2016, 14:2589-2595. |
| Gong L, Gan X Y, Zhang L, Chen Y C, Nie F J, Shi L, Guo Z Q, Song Y X. Cloning and function analysis of the StNAC72 gene from potato(Solanum tuberosum). Mol Plant Breed, 2016, 14:2589-2595 (in Chinese with English abstract). | |
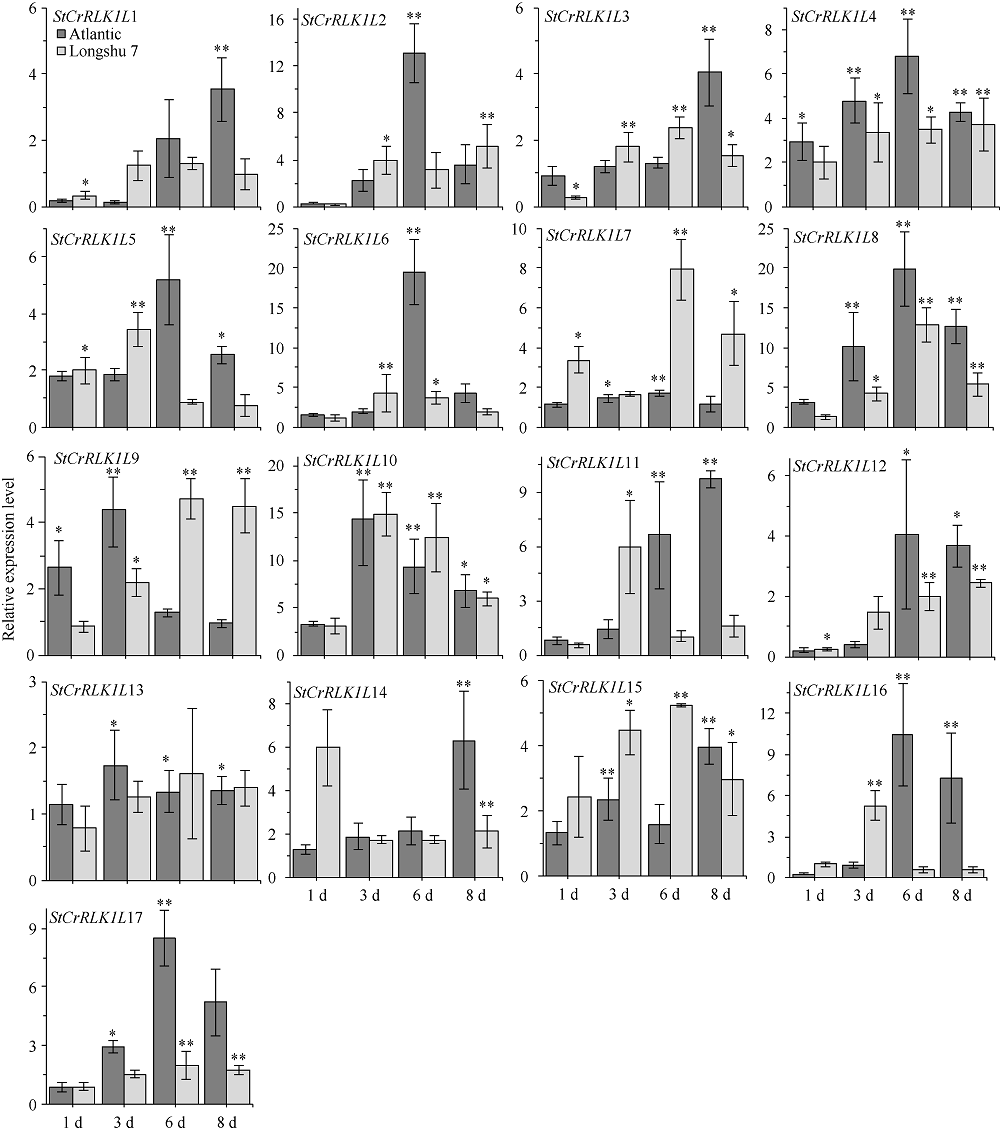
| [26] | Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-Time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25:402-408. |
| [27] | Hématy K, Sado P E, Van Tuinen A, Rochange S, Desnos T, Balzergue S, Pelletier S, Renou J P, Höfte H. A receptor-like kinase mediates the response of Arabidopsis cells to the inhibition of cellulose synthesis. Curr Biol, 2007, 17:922-931. |
| [28] | Yin Z, Ke X, Kang Z, Huang L. Apple resistance responses against valsa mali, revealed by transcriptomics analyses. Physiol Mol Plant, 2016, 93:85-92. |
| [29] | Niu E, Cai C, Zheng Y, Shang X, Fang L, Guo W. Genome-wide analysis of CrRLK1L gene family in Gossypium and identification of candidate CrRLK1L genes related to fiber development. Mol Genet Genomics, 2016, 291:1137-1154. |
| [30] | Liu P, Wan J, Guo Y, Ge S, Rao G. Adaptive evolution of the chrysanthemyl diphosphate synthase gene involved in irregular monoterpene metabolism. BMC Evol Biol, 2012, 12:214. |
| [31] | Zhang S. Evolutionary expansion, gene structure, and expression of the rice wall-associated kinase gene family. Plant Physiol, 2005, 139:1107-1124. |
| [32] | Xu G, Guo C, Shan H, Kong H. Divergence of duplicate genes in exon-intron structure. Proc Natl Acad Sci USA, 2012, 109:1187-1192. |
| [33] | Bari R, Jones J D. Role of plant hormones in plant defense response. Plant Mol Biol, 2009, 69:473-488. |
| [34] | Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol, 2005, 43:205-227. |
| [35] | Ballester A R, Norelli J, Burchard E, Abdelfattah A, Levin E, Gonzálezcandelas L, Droby S, Wisniewski M. Transcriptomic response of resistant (pi613981-pi613981-Malus sieversii) and susceptible (“royal gala”) genotypes of apple to blue mold (Penicillium expansum) infection. Front Plant Sci, 2017, 8:1981. |
| [36] | Solis-Miranda J, Fonseca-García C, Nava N, Pacheco R, Quinto C. Genome-wide identification of the CrRLK1L subfamily and comparative analysis of its role in the legume-rhizobia symbiosis. Genes, 2020, 11:793. |
| [1] | JIAN Hong-Ju, SHANG Li-Na, JIN Zhong-Hui, DING Yi, LI Yan, WANG Ji-Chun, HU Bai-Geng, Vadim Khassanov, LYU Dian-Qiu. Genome-wide identification and characterization of PIF genes and their response to high temperature stress in potato [J]. Acta Agronomica Sinica, 2022, 48(1): 86-98. |
| [2] | Wei-Na ZHANG,Yan-Ling FAN,Yi-Chen KANG,Xin-Yu YANG,Ming-Fu SHI,Kai YAO,Zhang-Ping ZHAO,Jun-Lian ZHANG,Shu-Hao QIN. Genome wide identification and expression analysis of CRK gene family in response to fungal pathogen signals in potato [J]. Acta Agronomica Sinica, 2020, 46(5): 680-689. |
|
||