欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (6): 914-923.doi: 10.3724/SP.J.1006.2020.94141
郭学民( ),赵晓曼,徐珂,王芯蕊,张辰瑜,东方阳
),赵晓曼,徐珂,王芯蕊,张辰瑜,东方阳
GUO Xue-Min(),ZHAO Xiao-Man,XU Ke,WANG Xin-Rui,ZHANG Chen-Yu,DONG-FANG Yang
摘要:
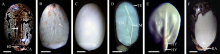
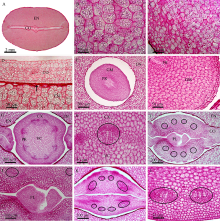
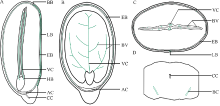
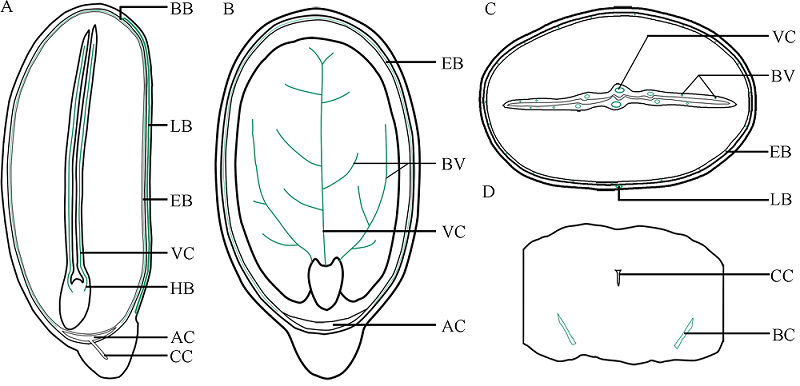
本文通过萌发法、分离法、石蜡切片法和显微观察技术, 分别以番红固绿和希夫试剂染色, 系统观察了蓖麻种皮、胚乳和胚的结构及其维管束分布。(1)蓖麻种子背侧基部的种孔并未被种阜所覆盖。(2)种皮包括外种皮、内种皮和种阜3部分, 其中, 外种皮由外至内分别为长柱状表皮层、海绵组织层和栅栏组织层, 而内种皮则依次为马氏层、海绵组织层和内珠被内层, 在种阜端由内种皮内层和外层共同围成气室, 类似于鸡蛋的气室。(3)在外种皮和内种皮中均有维管束分布, 其中外种皮的大型维管束仅分布在种子腹侧种脊的海绵组织层内, 它从种脐延伸至种阜相对一端; 而内种皮维管束也分布在海绵组织层中, 通过维管束索, 逐级分枝, 止于种阜端气室边缘; 内、外种皮的维管束通过合点端的连接点相连续, 共同构成种皮维管系统。(4)种阜由表皮和薄壁组织构成, 其中薄壁组织由外侧小型薄壁细胞和内侧大型薄壁细胞构成。种阜维管束仅2束, 分布在种阜中部腹侧, 独立构成种阜维管系统; 在种阜中, 还有种阜管道, 其内端开口于内种皮的气室, 外端为盲端。(5)胚乳由含大量糊粉粒的薄壁细胞构成, 其中未见维管束分布。在胚中, 维管束为外韧维管束, 它们分化于胚轴, 从胚轴顶端两侧分别进入2片子叶主脉, 在主脉中由4个逐步减少为1个, 在子叶中分枝变细, 构成胚维管系统; 子叶表皮和叶肉细胞中均含糊粉粒, 但在胚芽、胚轴和胚根中未见糊粉粒; 2片子叶近轴面分离, 远轴面则通过粘液层与胚乳相连接; 胚轴和胚根在空间上与胚乳分离。这些结果为全面掌握植物蓖麻种子的结构和研究种子发育和萌发过程及其物质运输提供了解剖学依据。
| [1] | Kadri A, Gharsallah N, Damak M, Gdoura R . Chemical composition and in vitro antioxidant properties of essential oil of Ricinus communis L. J Med Plants Res, 2011,5:1466-1470. |
| [2] | Zarai Z, Ben Chobba I, Ben Mansour R, Békir A, Gharsallah N, Kadri A . Essential oil of the leaves of Ricinus communis L.: In vitro cytotoxicity and antimicrobial properties. Lipids Health Disease, 2012,11:102. |
| [3] | Severino L S, Auld D L, Baldanzi M, Cândido M J D, Chen G, Crosby W, Tan D, He X, Lakshmamma P, Lavanya C, Machado O L T, Mielke T, Milani M, Miller T D, Morris J B, Morse S A, Navas A A, Soares D J, Sofiatti V, Wang M L, Zanotto M D, Zieler H . A review on the challenges for increased production of castor. Agron J, 2012,104:853-880. |
| [4] | Pius C O, Nnaemeka S P O, Charles O, Vincent N O, Chinenye A I, . Design enhancement evaluation of a castor seed shelling machine. J Sci Res Rep, 2014,3:924-938. |
| [5] | Rahmati H, Salehi S, Malekpour A, Farhangi F . Antimicrobial activity of castor oil plant ( Ricinus communis) seeds extract against gram positive bacteria, gram negative bacteria and yeast. Int J Mol Med Adv Sci, 2015,11:9-12. |
| [6] | Scarpa A, Guerci A . Various uses of the castor oil plant ( Ricinus communis L.): a review. J Ethnopharmacol, 1982,5:117-137. |
| [7] | Wafa G, Amadou D, Larbi K M, Héla E F O . Larvicidal activity, phytochemical composition, and antioxidant properties of different parts of five populations of Ricinus communis L. Ind Crop Prod, 2014,56:43-51. |
| [8] | Kaushik N K, Bagavan A, Rahuman A A, Zahir A A, Kamaraj C, Elango G, Jayaseelan C, Kirthi A V, Santhoshkumar T, Marimuthu S, Rajakumar G, Tiwari S K, Saha D . Evaluation of antiplasmodial activity of medicinal plants from North Indian Buchpora and South Indian Eastern Ghats. Malaria J, 2015, doi 10.1186/s12936-015-0564-z. |
| [9] | 曾佑炜, 宋光泉, 彭永宏, 梁关生 . 蓖麻毒蛋白研究及应用进展(综述). 亚热带植物科学, 2004, ( 1):60-63. |
| Zeng Y W, Song G Q, Peng Y H, Liang G S . A review of application and research progress in ricin. Subtrop Plant Sci, 2004, ( 1):60-63 (in Chinese with English abstract). | |
| [10] | 熊冬梅, 苏小军, 熊兴耀 . 蓖麻毒蛋白的提取及应用研究. 化学与生物工程, 2013,30(12):59-62. |
| Xiong D M, Su X J, Xiong X Y . Extraction and application research of ricin. Chem Bioengineer, 2013,30(12):59-62 (in Chinese with English abstract). | |
| [11] | 聶伟, 殷切, 刘小华 . 蓖麻毒蛋白作用的研究进展. 中国畜牧兽医文摘, 2016,32(10):80. |
| Nie W, Yin Q, Liu X H . Research progress on ricin. China Anim Husbandry Veterinary Med, 2016,32(10):80 (in Chinese). | |
| [12] | 张艳欣, 梁塔娜, 李丽丽, 杨丽凤, 常如慧, 姜桐桐, 孟祥越, 黄凤兰 . 蓖麻种子应用的研究进展. 贵州农业科学, 2018,46(5):12-15. |
| Zhang Y X, Liang T N, Li L L, Yang L F, Chang R H, Jiang T T, Meng X Y, Huang F L . Research progress in application of Ricinus communis seeds. Guizhou Agric Sci, 2018,46(5):12-15 (in Chinese with English abstract). | |
| [13] | Bianchini M, Pacini E . The caruncle of Ricinus communis L.( castor bean): its development and role in seed dehydration, rehydration, and germination. Int J Plant Sci, 1996,157:40-48. |
| [14] | Lisci M, Bianchini M, Pacini E . Structure and function of the elaiosome in some angiosperm species. Flora, 1996,191:131-141. |
| [15] | Kriedemann P, Beevers H . Sugar uptake and translocation in the castor bean seedling: I. Characteristics of transfer in intact and excised Seedlings. Plant Physiol, 1967,42:161-173. |
| [16] | Vigil E L . Cytochemical and developmental changes in microbodies (glyoxysomes) and related organelles of castor bean endosperm. J Cell Biol, 1970,46:435-454. |
| [17] | Fabbri F, Palandri M . Ultrastructural modifications in contyledonous leaves of Ricinus comminis L. during aging. Caryologia, 1970,23:677-714. |
| [18] | 陆时万, 徐祥生, 沈敏健 . 植物学 (第2版上册). 北京: 高等教育出版社, 1992. pp 35, 70. |
| Lu S W, Xu X S, Shen M J. Botany, 2nd edn. Part I. Beijing: Higher Education Press, 1992. pp 35, 70 (in Chinese). | |
| [19] | 马炜梁 . 植物学. 北京: 高等教育出版社, 2012. pp 24, 101. |
| Ma W L. Botany. Beijing: Higher Education Press, 2012. pp 24, 101 (in Chinese). | |
| [20] | 郭学民, 徐兴友, 孟宪东, 左照江, 高荣孚 . 合欢种子硬实与萌发特性及种皮微形态与结构特征的研究. 内蒙古农业大学学报(自然科学版), 2006,27(3):13-18. |
| Guo X M, Xu X Y, Meng X D, Zuo Z J, Gao R F . Bourgeon characteristics of hardseed of Albizia julibrissin Durazz. and its testa micro-morphology and structure. J Inner Mongolia Agric Univ( Nat Sci Edn), 2006,27(3):13-18 (in Chinese with English abstract). | |
| [21] | Sernander R . Entwurf einer Monographie der europäischen Myrmekochoren. Uppsala: Almqvist & Wiksells, 1906 (in German). |
| [22] | Bhojwani S S, Bhantnagar S P. The Embryology of Angiosperms3rd edn. Delhi: Vikas Publishing House PVT Ltd, 1979. p 151. |
| [23] | 王凯基, 倪德祥 . 植物生物学词典. 上海: 上海科技教育出版社, 1998. p 378. |
| Wang K J, Ni D X. Dictionary of Plant Biology. Shanghai: Shanghai Science and Technology Education Press, 1998. p 378 (in Chinese). | |
| [24] | Pacini E . Mercurialis annua L. (Euphorbiaceae) seed interacyions with the ant Messor strutor (Latr.). Hymenoptera: Formicidae. Acta Bot Neerl, 1990,39:253-262. |
| [25] | Kriedemann P, Beevers H . Sugar uptake and translocation in the castor bean seedling: II. Sugar transformations during uptake. Plant Physiol, 1967,42:174-180. |
| [26] |
Komor E . Sucrose uptake by cotyledons of Ricinus communis L.: characteristics, mechanism, and regulation. Planta, 1977,137:119-131.
doi: 10.1007/BF00387548 |
| [1] | 雷新慧, 万晨茜, 陶金才, 冷佳俊, 吴怡欣, 王家乐, 王鹏科, 杨清华, 冯佰利, 高金锋. 褪黑素与2,4-表油菜素内酯浸种对盐胁迫下荞麦发芽与幼苗生长的促进效应[J]. 作物学报, 2022, 48(5): 1210-1221. |
| [2] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [3] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [4] | 李振华, 王显亚, 刘一灵, 赵杰宏. NtPHYB1与光温信号互作调控烟草种子萌发[J]. 作物学报, 2022, 48(1): 99-107. |
| [5] | 程艳双, 胡美艳, 杜志敏, 闫秉春, 李丽, 王祎玮, 鞠晓堂, 孙丽丽, 徐海. 减氮对辽粳5号/秋田小町RIL群体茎秆维管束、穗部和产量 性状的影响及其相互关系[J]. 作物学报, 2021, 47(5): 964-973. |
| [6] | 吕冬梅, 朱广龙, 王玥, 施雨, 卢发光, 任桢, 刘昱茜, 顾立峰, 卢海潼, Irshad Ahmad, 焦秀荣, 孟天瑶, 周桂生. 苗期重金属胁迫下蓖麻生长、生理和重金属积累效应[J]. 作物学报, 2021, 47(4): 728-737. |
| [7] | 唐婧泉, 王南, 高界, 刘婷婷, 文静, 易斌, 涂金星, 傅廷栋, 沈金雄. 甘蓝型油菜SnRK基因家族生物信息学分析及其与种子含油量的关系[J]. 作物学报, 2021, 47(3): 416-426. |
| [8] | 邱红梅, 陈亮, 侯云龙, 王新风, 陈健, 马晓萍, 崔正果, 张玲, 胡金海, 王跃强, 邱丽娟. 大豆种子颜色遗传调控机制研究进展[J]. 作物学报, 2021, 47(12): 2299-2313. |
| [9] | 孟鑫浩, 张靖男, 崔顺立, Charles Y.Chen, 穆国俊, 侯名语, 杨鑫雷, 刘立峰. 花生荚果与种子相关性状QTL定位及与环境互作分析[J]. 作物学报, 2021, 47(10): 1874-1890. |
| [10] | 鲁庚,唐鑫,陆俊杏,李丹,胡秋芸,胡田,张涛. 紫苏二酰基甘油酰基转移酶2基因克隆与功能研究[J]. 作物学报, 2020, 46(8): 1283-1290. |
| [11] | 晁毛妮,胡海燕,王润豪,陈煜,付丽娜,刘庆庆,王清连. 陆地棉钾转运体基因GhHAK5启动子的克隆与功能分析[J]. 作物学报, 2020, 46(01): 40-51. |
| [12] | 张双双,王立伟,姚楠,郭光艳,夏玉凤,秘彩莉. 水稻OsUBA基因的表达及其在促进种子萌发和开花中的功能[J]. 作物学报, 2019, 45(9): 1327-1337. |
| [13] | 宋松泉,刘军,徐恒恒,张琪,黄荟,伍贤进. 乙烯的生物合成与信号及其对种子萌发和休眠的调控[J]. 作物学报, 2019, 45(7): 969-981. |
| [14] | 崔月,陆建农,施玉珍,殷学贵,张启好. 蓖麻株高性状主基因+多基因遗传分析[J]. 作物学报, 2019, 45(7): 1111-1118. |
| [15] | 申状状,李昱樱,荣二花,吴玉香. 陆地棉和野生斯特提棉种间异源六倍体的合成与性状鉴定[J]. 作物学报, 2019, 45(4): 628-634. |
|
||