欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (5): 837-846.doi: 10.3724/SP.J.1006.2021.04173
马贵芳1( ), 满夏夏1, 张益娟2, 高豪1, 孙朝霞1,3, 李红英1,3, 韩渊怀1,3, 侯思宇1,3,*()
), 满夏夏1, 张益娟2, 高豪1, 孙朝霞1,3, 李红英1,3, 韩渊怀1,3, 侯思宇1,3,*()
MA Gui-Fang1(), MAN Xia-Xia1, ZHANG Yi-Juan2, GAO Hao1, SUN Zhao-Xia1,3, LI Hong-Ying1,3, HAN Yuan-Huai1,3, HOU Si-Yu1,3,*()
摘要:
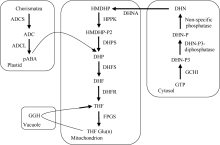
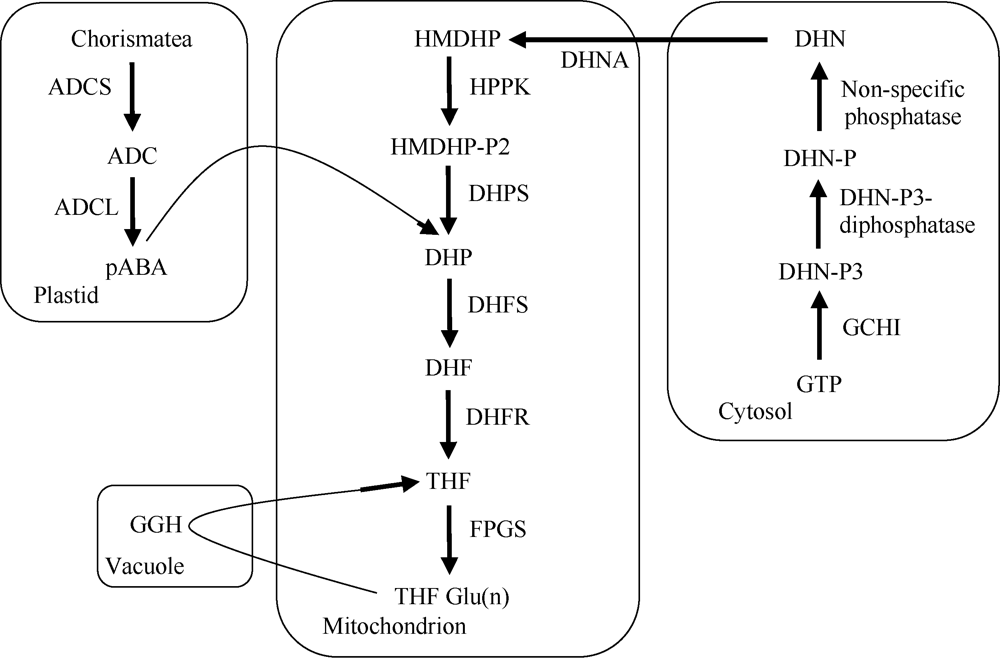
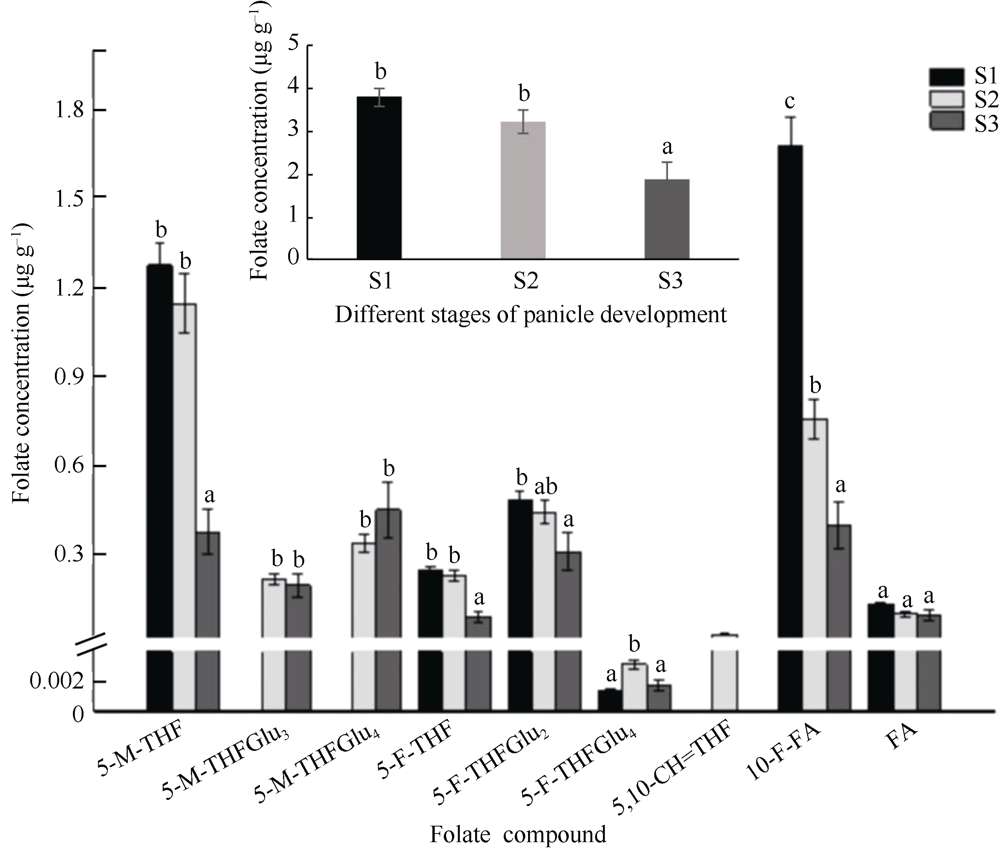
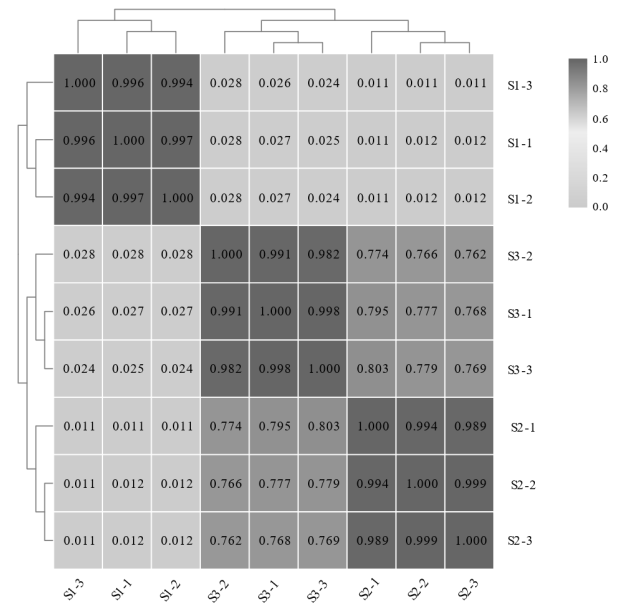
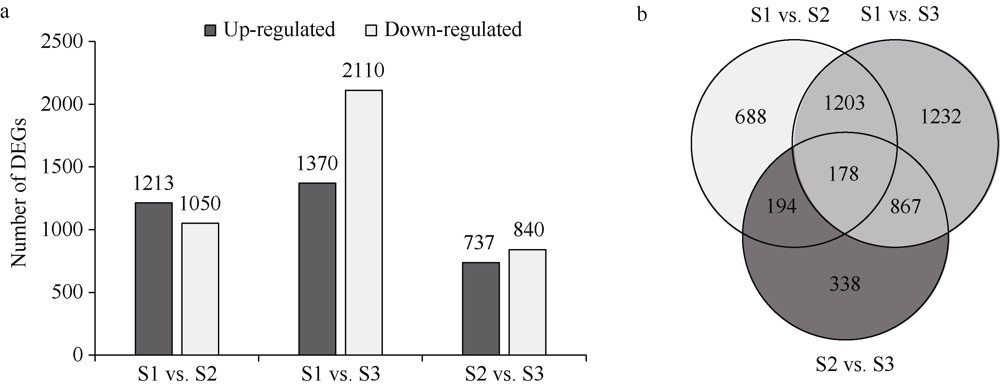
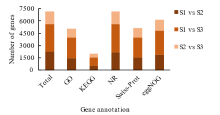
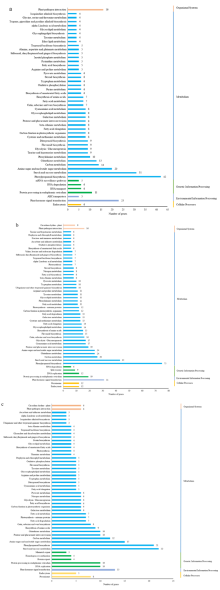
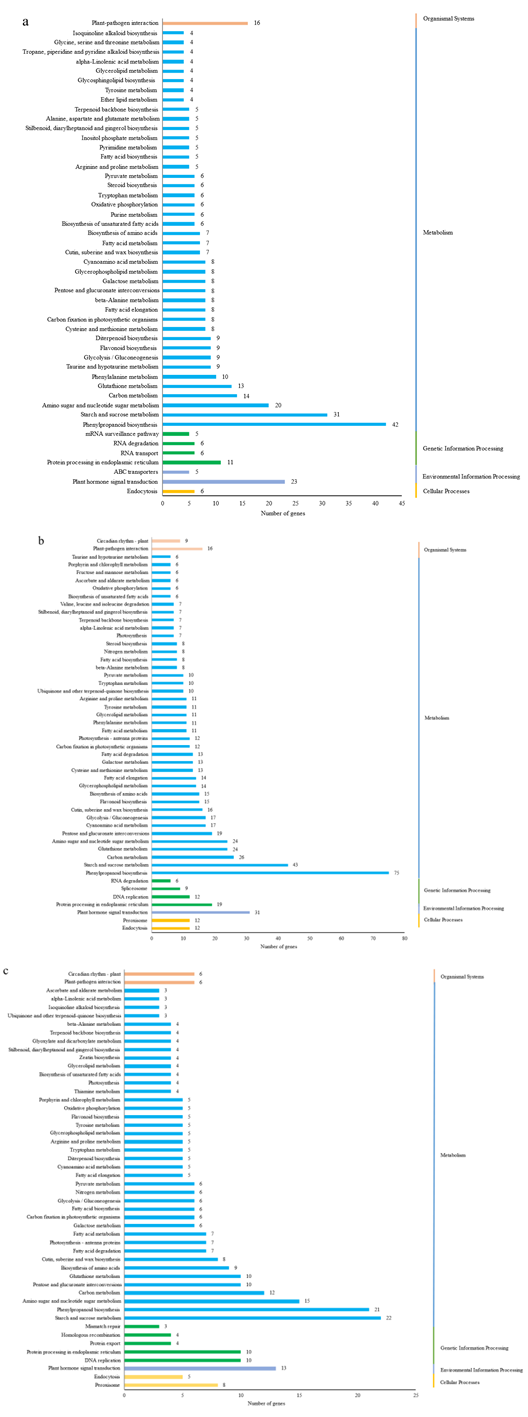
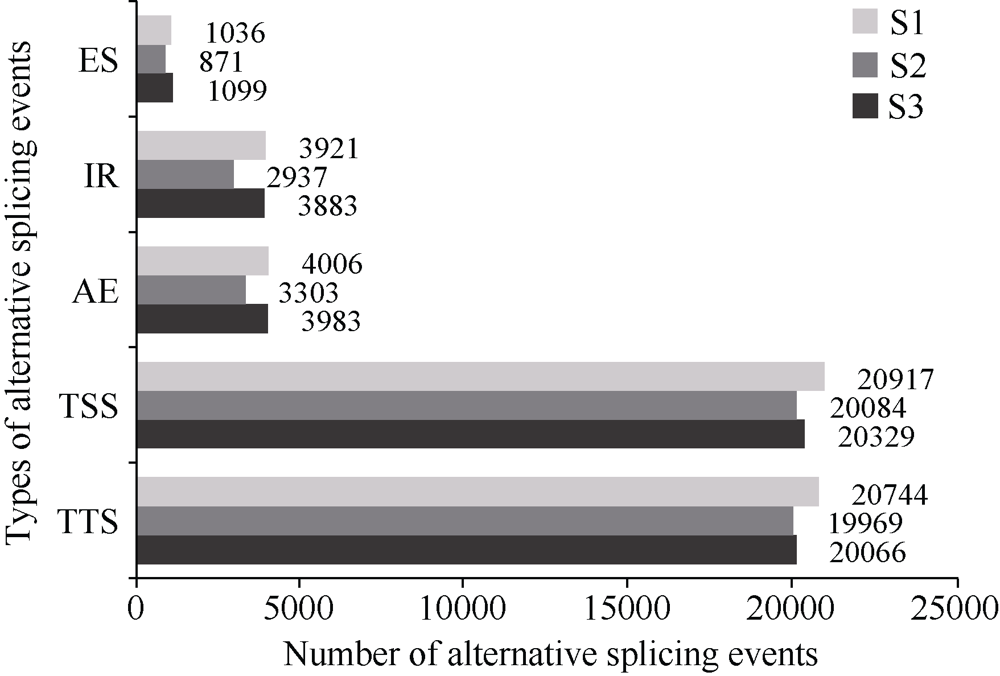
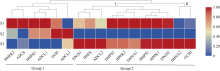
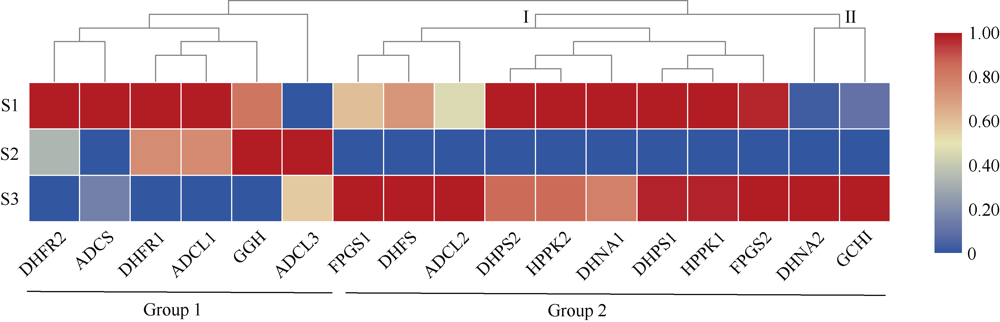
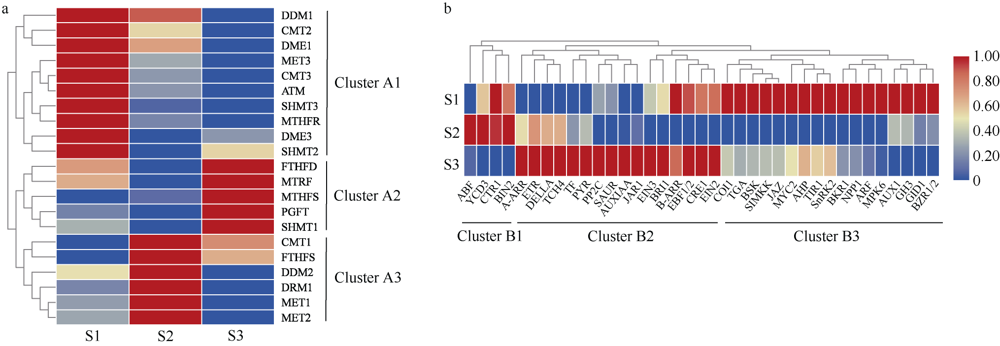
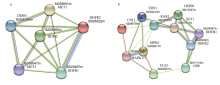
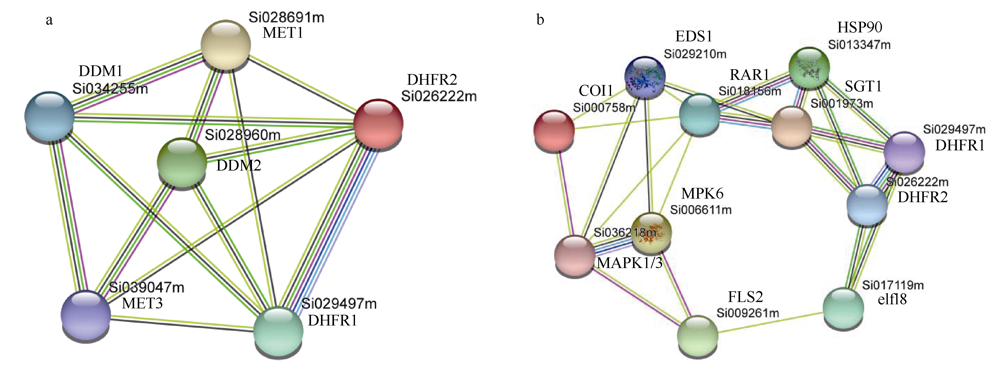
植物叶酸(Folate, FA)参与细胞内一碳单位转移反应, 是能量代谢、氨基酸和核酸合成的重要供体。前期我们发现, 谷子籽粒叶酸含量高于其他禾谷类作物, 但叶酸代谢组分特征目前尚不清楚。本研究以‘晋谷21’为试验材料, 对3个穗发育期小穗进行叶酸代谢组分析; 通过RNA-seq分析穗发育期差异表达基因及叶酸合成途径相关代谢通路, 探究叶酸组分与叶酸代谢途径基因表达模式及其与可变剪切的相关性, 同时预测分析叶酸合成途径基因的蛋白互作网络。结果表明, 总叶酸含量随穗发育阶段呈下降趋势; 5-甲基四氢叶酸(5-methyltetrahydrofolate, 5-M-THF)和10-甲酰叶酸(10-formyl folic acid, 10-F-FA)是穗发育期的主要组分。17个叶酸合成关键基因表达模式可划分为2个不同类群。转录起始区域(transcription start site, TSS)和转录终止区域(transcription terminal site, TTS)可变剪切是叶酸合成关键基因在穗发育3个时期均发生的可变剪切类型, 外显子跳跃型(exon skipping, ES)、内含子滞留型(intron retention, IR)和可变5'或3'端剪切(alternative exon ends, AE)是特有类型。同时发现叶酸合成、一碳代谢和激素信号传导途径关键基因与叶酸组分差异具有相关性, 初步推测谷子穗发育过程叶酸合成途径及偶联途径相关基因的表达一定程度影响叶酸含量变化, 其中ADCS、DHFR2和GGH基因的差异表达可能为影响谷穗叶酸含量的主要原因, 可作为利用遗传工程提高谷子叶酸含量及生物强化分子育种的关键靶基因。
| [1] |
Young D W. The biosynthesis of the vitamins thiamin, riboflavin, and folic acid. Nat Prod Rep, 1986,3:395-419.
doi: 10.1039/np9860300395 pmid: 3547188 |
| [2] |
Chistoserdova L, Vorholt J A, Thauer R K, Lidstrom M E. C1 transfer enzymes and coenzymes linking methylotrophic bacteria and methanogenic Archaea. Science, 1998,281:99-102.
doi: 10.1126/science.281.5373.99 pmid: 9651254 |
| [3] | 胡廷章. 生物体中一碳单位的代谢. 四川三峡学院学报, 1999, (5):74-77. |
| Hu Y Z. The metabolism of one carbon uni tin living. J Chongqing Three Gorges Univ, 1999, (5):74-77 (in Chinese with English abstract). | |
| [4] | Strum W, Nixon P F, Bertino J B, Bertino H J. Intestinal folate absorption: I. 5-methyltetrahydrofolic acid. J Investig, 1971,50:1910-1916. |
| [5] |
Shane B. Folylpolyglutamate synthesis and role in the regulation of one-carbon metabolism. Vitam Horm, 1989,45:263-335.
doi: 10.1016/s0083-6729(08)60397-0 pmid: 2688305 |
| [6] | 单齐冀. 糯玉米种质资源评价及籽粒发育过程叶酸合成代谢规律研究. 华南理工大学硕士学位论文, 广东广州, 2019. |
| Shan Q J. The Germplasm Evaluation and Folate Synthetic Metabolism during Kernel Development of Waxy Maize. MS Thesis of South China University of Technology, Guangzhou, Guangdong, China, 2019 (in Chinese with English abstract). | |
| [7] |
Gorelova V, Bastien O, De Clerck O, Lespinats S, Rébeillé F, Van Der Straeten D. Evolution of folate biosynthesis and metabolism across algae and land plant lineages. Sci Rep, 2019,9:99-102.
doi: 10.1038/s41598-018-37471-0 pmid: 30643172 |
| [8] | 张大众, 刘佳佳, 冯佰利. 中国谷子种植利用史及其演进启示. 草业学报, 2018,27(3):173-186. |
| Zhang D Z, Liu J J, Feng B L. History of foxtail millet planting, husbandry, and societal utilization in China. Acta Pratacult Sin, 2018,27(3):173-186 (in Chinese with English abstract). | |
| [9] | 郑楠楠, 綦文涛, 王春玲, 贠婷婷, 幺杨, 任贵兴. 不同品种谷子营养成分及功能活性成分差异化分析. 粮油食品科技, 2018,26(2):34-39. |
| Zheng N N, Qi W T, Wang C L, Yun T T, Yao Y, Ren G X. Comparative analysis of nutritional and functional components in different kinds of millet. Sci Technol Cereals Oils Foods, 2018,26(2):34-39 (in Chinese with English abstract). | |
| [10] | Paul M F. Dietary Reference intakes for thiamin, riboflavin, niacin, vitamin B6, folate, vitamin B12, pantothenic acid, biotin and choline. Trends Food Sci Technol, 2000,11:296-297. |
| [11] | 邵丽华. 山西省小米叶酸含量的研究. 山西师范大学硕士学位论文, 山西临汾, 2014. |
| Shao L H. Study of Folic Acid Content in Millet of Shanxi Province. MS Thesis of Shanxi Normal University, Linfen, Shanxi, China, 2014 (in Chinese with English abstract). | |
| [12] | 侯思宇, 宋敏, 闫陆飞, 孙朝霞, 韩渊怀, 李红英. HPLC法测定谷子籽粒叶酸含量及种质资源评价. 土壤, 2018,50:1235-1240. |
| Hou S Y, Song M, Yan L F, Sun Z X, Han Y H, Li H Y. Evaluation and detection of folic acid content of seeds among foxtail millet accessions by HPLC method. Soils, 2018,50:1235-1240 (in Chinese with English abstract). | |
| [13] | Blancquaert D, Van Daele J, Strobbe S, Kiekens F, Storozhenko S, De Steur H, Gellynck X, Lambert W, Stove C, Van Der Straeten D. Improving folate (vitamin B9) stability in biofortified rice through metabolic engineering. Nat Biotechnol, 2015,33:1076-1078. |
| [14] |
Jacob J A. Twenty years after folic acid fortification, FDA ponders expansion to corn masa flour. JAMA, 2016,315:1821-1822.
doi: 10.1001/jama.2016.2611 pmid: 27082432 |
| [15] |
Mahato A, Vyas S, Chatterjee N S. HPLC-UV estimation of folic acid in fortified rice and wheat flour using enzymatic extraction and immunoaffinity chromatography enrichment: an interlaboratory validation study. J AOAC Int, 2020,103:73-77.
doi: 10.5740/jaoacint.19-0207 pmid: 31455468 |
| [16] |
Storozhenko S, Brouwer D V, Volckaert M, Navarrete O, Blancquaert D, Zhang G F, Lambert W, Van Der Straeten D. Folate fortification of rice by metabolic engineering. Nat Biotechnol, 2007,25:1277-1279.
doi: 10.1038/nbt1351 pmid: 17934451 |
| [17] | 汪冉冉. 过量表达GCHI和ADCS基因对提高植物叶酸含量的研究. 兰州大学硕士学位论文, 甘肃兰州, 2013. |
| Wang R R. The Study on Enhancing Folate Content in Plants by Overexpression of GCHI and ADCS Genes. MS Thesis of Lanzhou University, Lanzhou, Gansu, China, 2013 (in Chinese with English abstract). | |
| [18] | Wan X, Han L D, Yang M, Zhang H Y, Zhang C Y, Hu P. Simultaneous extraction and determination of mono-polyglutamyl folates using high-performance liquid chromatography-tandem mass spectrometry and its applications in starchy crops. Analy Bioanaly Chem, 2019,411:2891-2904. |
| [19] | Lian T, Guo W Z, Chen M R, Li J L, Liang Q J, Liu F, Meng H Y, Xu B S, Chen J F, Zhang C Y, Jiang L. Genome-wide identification and transcriptional analysis of folate metabolism-related genes in maize kernels. BMC Plant Biol, 2015,15:1-14. |
| [20] | 韩娟英, 何曦, 蒋宙蕾, 梅沙, 张宁, 吴殿星. 富含叶酸水稻研究进展. 中国稻米, 2017,23(6):7-12. |
| Han J Y, He X, Jiang Z L, Mei S, Zhang N, Wu D X. Progress on high folate content rice. China Rice, 2017,23(6):7-12 (in Chinese with English abstract). | |
| [21] |
Blancquaert D, Van Daele J, Storozhenko S, Stove C P, Lambert W, Van Der Straeten D. Rice folate enhancement through metabolic engineering has an impact on rice seed metabolism, but does not affect the expression of the endogenous folate biosynthesis genes. Plant Mol Biol, 2013,83:329-349.
doi: 10.1007/s11103-013-0091-7 pmid: 23771598 |
| [22] |
Hanson A D, Gregory J F. Folate biosynthesis, turnover, and transport in plants. Annu Rev Plant Biol, 2011,62:105-125.
doi: 10.1146/annurev-arplant-042110-103819 pmid: 21275646 |
| [23] |
Albani D, Parisi B, Carbonera D, Cella R. Dihydrofolate reductase from Daucus carota cell suspension cultures: purification, molecular and kinetic characterization. Plant Mol Biol, 1985,5:363-372.
pmid: 24306990 |
| [24] |
Bachmann B, Follmann H. Deoxyribonucleotide biosynthesis in green algae: characterization of thymidylate synthase-dihydrofolate reductase in Scenedesmus obliquus. Arch Biochem Biophys, 1987,256:244-252.
doi: 10.1016/0003-9861(87)90442-5 pmid: 3606123 |
| [25] |
Gorelova V, De Lepeleire J, Van Daele J, Pluim D, Meï C, Cuypers A, Leroux O, Rébeillé F, Schellens J H, Blancquaert D, Stove C P, Van Der Straeten D. Dihydrofolate reductase/ Thymidylate synthase fine-tunes the folate status and controls redox homeostasis in plants. Plant Cell, 2017,29:2831-2853.
doi: 10.1105/tpc.17.00433 pmid: 28939595 |
| [26] |
Kim S E, Cole P D, Cho R C, Ly A, Ishiguro L, Sohn K J, Croxford R, Kamen B A, Kim Y I. γ-glutamyl hydrolase modulation and folate influence chemosensitivity of cancer cells to 5-fluorouracil and methotrexate. British J Cancer, 2013,109:2175-2188.
doi: 10.1038/bjc.2013.579 |
| [27] |
Cheng C Y, Krishnakumar V, Chan A P, Thibaud N F, Schobel S, Town C D. Araport11: a complete reannotation of the Arabidopsis thaliana reference genome. Plant J, 2017,89:789-804.
doi: 10.1111/tpj.13415 pmid: 27862469 |
| [28] |
Swarup R, Crespi M, Bennett M J. One gene, many proteins: mapping cell-specific alternative splicing in plants. Dev Cell, 2016,39:383-385.
doi: 10.1016/j.devcel.2016.11.002 pmid: 27875679 |
| [29] |
Zhang X, Mount S M. Two alternatively spliced isoforms of the Arabidopsis SR45 protein have distinct roles during normal plant development. Plant Physiol, 2009,150:1450-1458.
doi: 10.1104/pp.109.138180 pmid: 19403727 |
| [30] |
Airoldi C A, Bergonzi S, Davies B. Single amino acid change alters the ability to specify male or female organ identity. Proc Natl Acad Sci USA, 2010,107:18898-18902.
doi: 10.1073/pnas.1009050107 pmid: 20956314 |
| [31] |
Severing E I, Dijk A D J V, Morabito G, Lange J B, Immink R G H, van Ham R C H J. Predicting the impact of alternative splicing on plant MADS domain protein function. PLoS One, 2012,7:e30524.
doi: 10.1371/journal.pone.0030524 pmid: 22295091 |
| [32] |
Chen M X, Zhang K L, Gao B, Yang J A, Tian Y, Das D, Fan D, Dai L, Hao G, Yang G, Zhang J H, Zhu F, Fang Y M. Phylogenetic comparison of 5′ splice site determination in central spliceosomal proteins of the U1-70K gene family, in response to developmental cues and stress conditions. Plant J, 2020,103:357-378.
doi: 10.1111/tpj.14735 pmid: 32133712 |
| [33] |
González B, Vera P. Folate metabolism interferes with plant immunity through 1C methionine synthase-directed genome-wide DNA methylation enhancement. Mol Plant, 2019,12:1227-1242.
pmid: 31077872 |
| [34] | John S, Fabrice R, John F. Folic acid and folates: the feasibility for nutritional enhancement in plant foods. J Sci Food Agric, 2000,80:795-824. |
| [35] |
Wang L, Kong D D, Lyu Q, Niu G Q, Han T T, Zhao X C, Meng S L, Cheng Q, Guo S C, Du J, Wu Z L, Wang J Z, Bao F, Hu Y, Pan X J, Xia J C, Yuan D, Han L D, Lian T, Zhang C Y, Wang H Y, He X J, He Y K. Tetrahydrofolate modulates floral transition through epigenetic silencing. Plant Physiol, 2017,174:1274-1284.
doi: 10.1104/pp.16.01750 pmid: 28450424 |
| [36] |
Finni W, Basem K, Marion W, Thomas H, Katrin F O, Wilfried S, Philippe S K, Corina V A. Folic acid induces salicylic acid-dependent immunity in Arabidopsis and enhances susceptibility to Alternaria brassicicola. Mol Plant Pathol, 2015,16:616-622.
doi: 10.1111/mpp.12216 pmid: 25348251 |
| [37] | 肖熙鸥, 林文秋, 李可, 冯雪峰, 李威, 邹华芬, 金辉. 茄子Enhanced disease susceptibility 1互作蛋白的初步筛选与分析. 热带作物学报, 2018,39:1580-1584. |
| Xiao X O, Lin W Q, Li K, Feng X F, Li W, Zou H F, Jin H. Screening and analysis of candidate interactors of enhanced disease susceptibility 1 (SmEDS1) in eggplant. Chin J Trop Crops, 2018,39:1580-1584 (in Chinese with English abstract). |
| [1] | 晋敏姗, 曲瑞芳, 李红英, 韩彦卿, 马芳芳, 韩渊怀, 邢国芳. 谷子糖转运蛋白基因SiSTPs的鉴定及其参与谷子抗逆胁迫响应的研究[J]. 作物学报, 2022, 48(4): 825-839. |
| [2] | 杜晓芬, 王智兰, 韩康妮, 连世超, 李禹欣, 张林义, 王军. 谷子叶绿体基因RNA编辑位点的鉴定与分析[J]. 作物学报, 2022, 48(4): 873-885. |
| [3] | 赵美丞, 刁现民. 谷子近缘野生种的亲缘关系及其利用研究[J]. 作物学报, 2022, 48(2): 267-279. |
| [4] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [5] | 汪颖, 高芳, 刘兆新, 赵继浩, 赖华江, 潘小怡, 毕晨, 李向东, 杨东清. 利用WGCNA鉴定花生主茎生长基因共表达模块[J]. 作物学报, 2021, 47(9): 1639-1653. |
| [6] | 曹亮, 杜昕, 于高波, 金喜军, 张明聪, 任春元, 王孟雪, 张玉先. 外源褪黑素对干旱胁迫下绥农26大豆鼓粒期叶片碳氮代谢调控的途径分析[J]. 作物学报, 2021, 47(9): 1779-1790. |
| [7] | 牛丽, 白文波, 李霞, 段凤莹, 侯鹏, 赵如浪, 王永宏, 赵明, 李少昆, 宋吉青, 周文彬. 地膜覆盖对黄土高原地区两种种植密度下玉米叶片代谢组的影响[J]. 作物学报, 2021, 47(8): 1551-1562. |
| [8] | 黄文功, 姜卫东, 姚玉波, 宋喜霞, 刘岩, 陈思, 赵东升, 吴广文, 袁红梅, 任传英, 孙中义, 吴建忠, 康庆华. 亚麻响应低钾胁迫转录谱分析[J]. 作物学报, 2021, 47(6): 1070-1081. |
| [9] | 李鹏程, 毕真真, 孙超, 秦天元, 梁文君, 王一好, 许德蓉, 刘玉汇, 张俊莲, 白江平. DNA甲基化参与调控马铃薯响应干旱胁迫的关键基因挖掘[J]. 作物学报, 2021, 47(4): 599-612. |
| [10] | 贾小平, 李剑峰, 张博, 全建章, 王永芳, 赵渊, 张小梅, 王振山, 桑璐曼, 董志平. 谷子SiPRR37基因对光温、非生物胁迫的响应特点及其有利等位变异鉴定[J]. 作物学报, 2021, 47(4): 638-649. |
| [11] | 王瑞莉, 王刘艳, 雷维, 吴家怡, 史红松, 李晨阳, 唐章林, 李加纳, 周清元, 崔翠. 结合RNA-seq分析和QTL定位筛选甘蓝型油菜萌发期与铝毒胁迫相关的候选基因[J]. 作物学报, 2021, 47(12): 2407-2422. |
| [12] | 张欢, 罗怀勇, 李威涛, 郭建斌, 陈伟刚, 周小静, 黄莉, 刘念, 晏立英, 雷永, 廖伯寿, 姜慧芳. 花生全基因组抗病基因鉴定及其对青枯菌侵染的响应分析[J]. 作物学报, 2021, 47(12): 2314-2323. |
| [13] | 曾健, 徐先超, 徐昱斐, 王秀成, 于海燕, 冯贝贝, 邢光南. 利用动态转录组学挖掘大豆百粒重候选基因[J]. 作物学报, 2021, 47(11): 2121-2133. |
| [14] | 秦天元, 孙超, 毕真真, 梁文君, 李鹏程, 张俊莲, 白江平. 基于WGCNA的马铃薯根系抗旱相关共表达模块鉴定和核心基因发掘[J]. 作物学报, 2020, 46(7): 1033-1051. |
| [15] | 贾小平,袁玺垒,李剑峰,王永芳,张小梅,张博,全建章,董志平. 不同光温条件谷子光温互作模式研究及SiCCT基因表达分析[J]. 作物学报, 2020, 46(7): 1052-1062. |
|
||