欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (1): 151-164.doi: 10.3724/SP.J.1006.2022.11005
马博闻( ), 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑*(), 姜东*()
), 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑*(), 姜东*()
MA Bo-Wen(), LI Qing, CAI Jian, ZHOU Qin, HUANG Mei, DAI Ting-Bo, WANG Xiao*(), JIANG Dong*()
摘要:
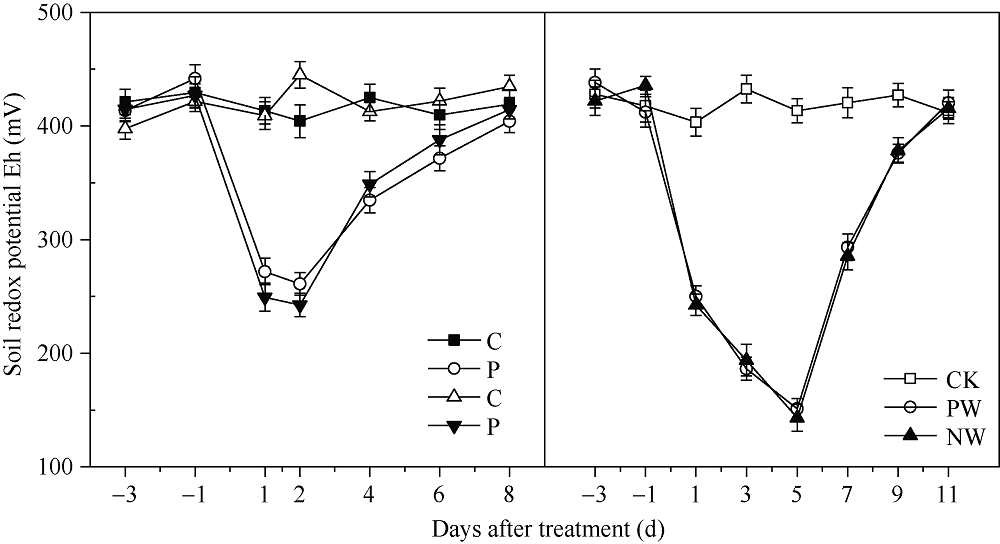
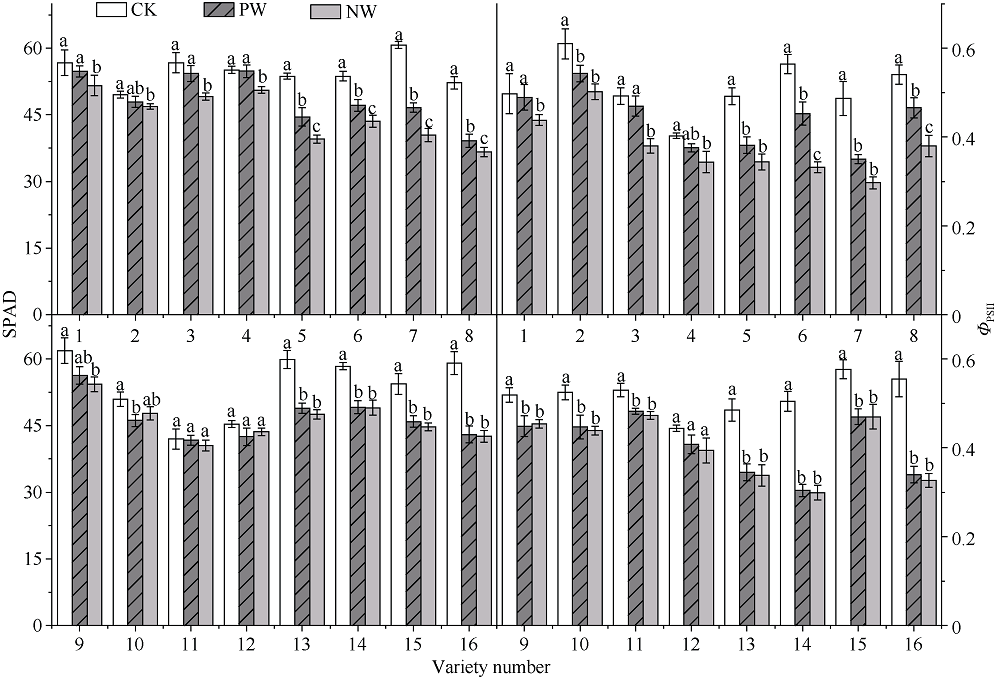
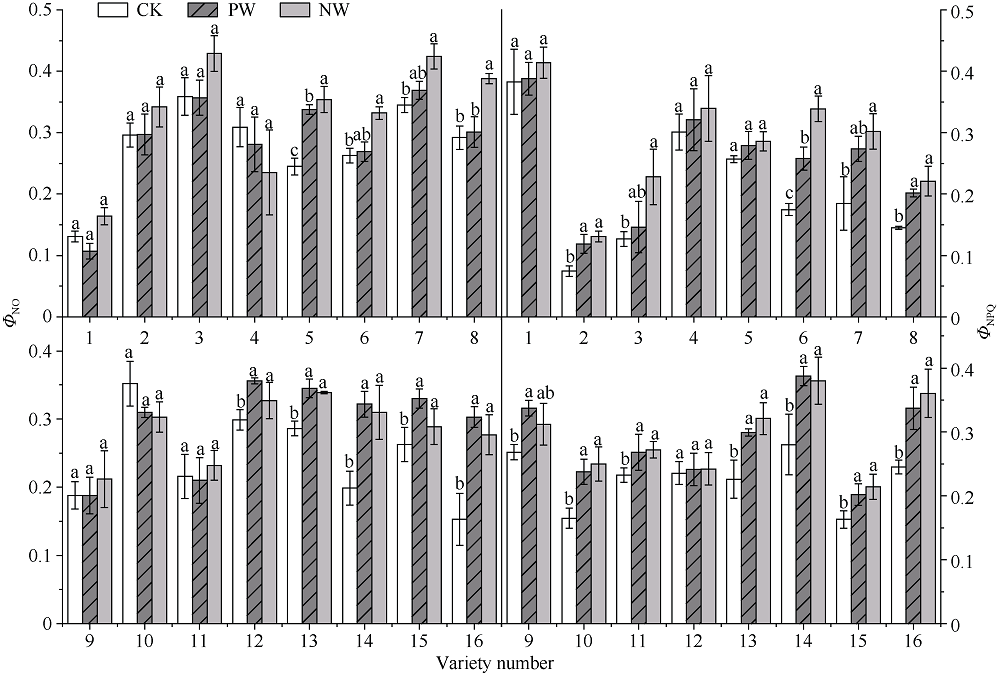
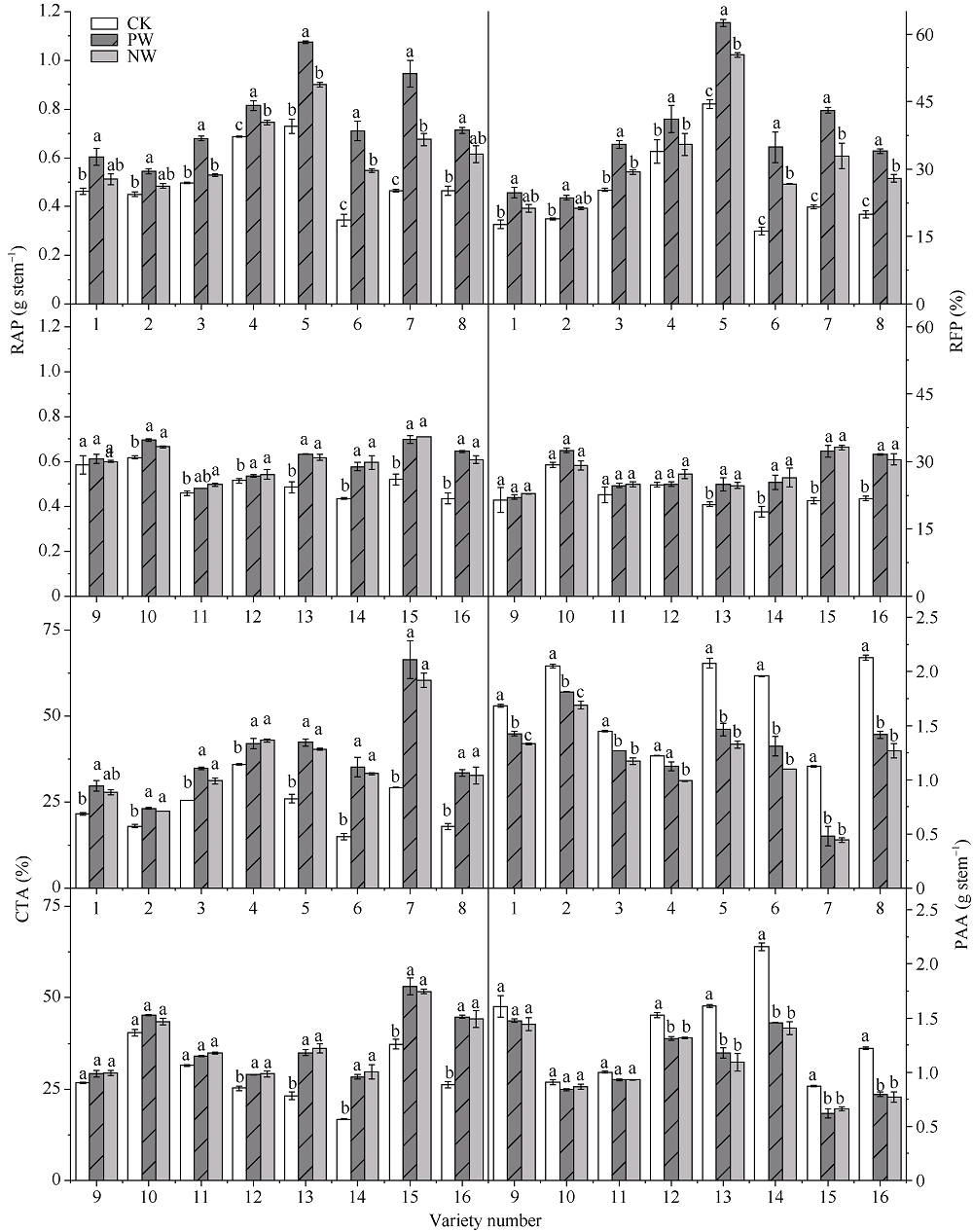
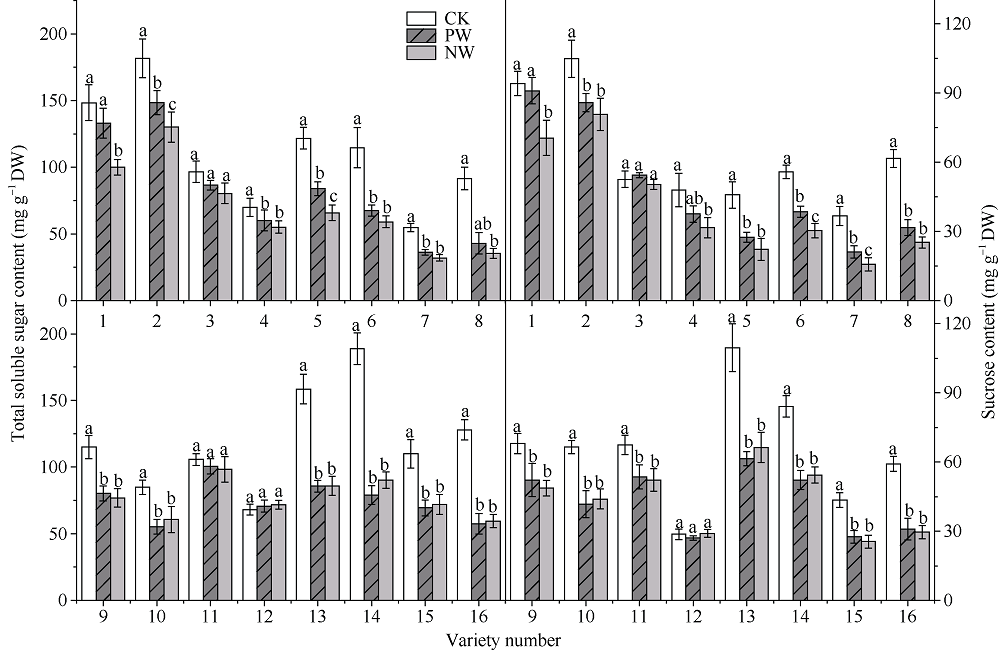
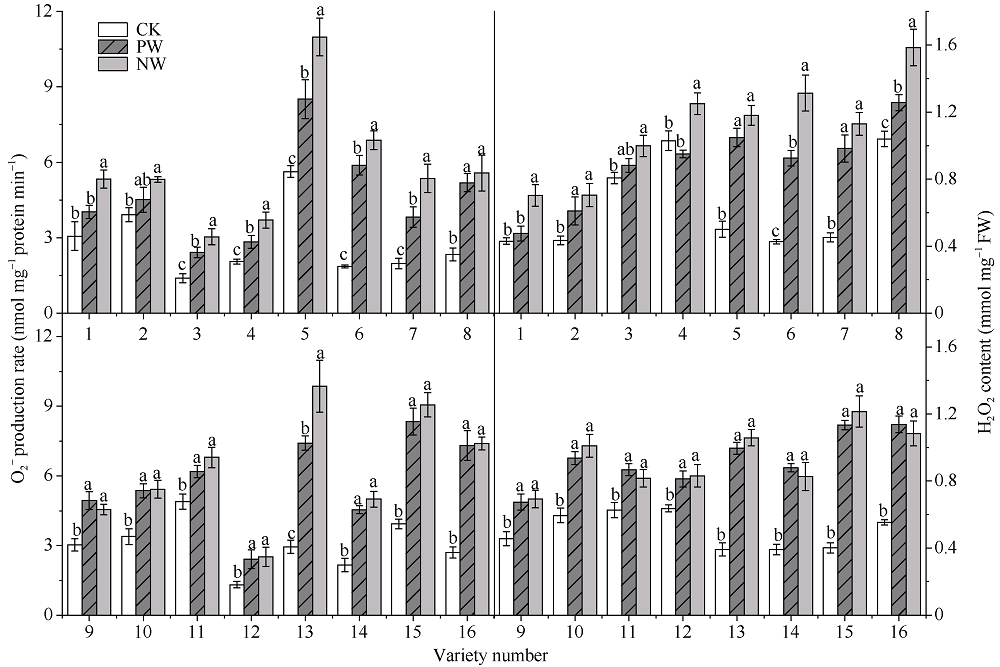
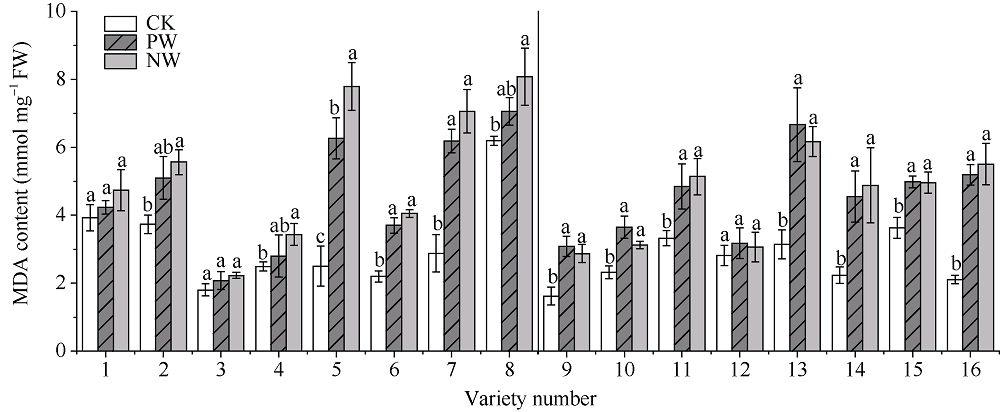
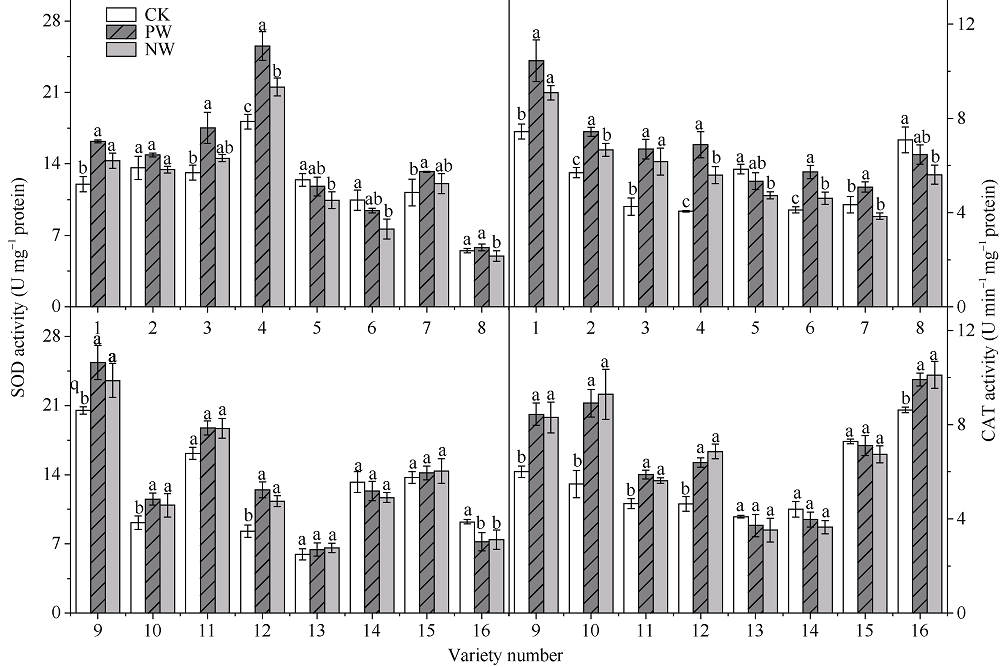
采用对渍水胁迫和渍水锻炼响应差异的小麦品种为材料, 在四叶一心期和六叶一心期分别进行渍水锻炼2 d; 在开花后7 d进行渍水胁迫5 d, 分析不同小麦品种对渍水胁迫响应的差异及其生理机制。结果表明, 花后渍水胁迫显著降低旗叶叶绿素含量(SPAD)和实际光化学效率(ΦPSII), 抑制花后光合同化物积累(PAA), 导致籽粒千粒重和产量降低; 与不耐渍型品种相比, 耐渍型品种在渍水胁迫下可维持较高的SPAD、ΦPSII和PAA, 超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)以及谷胱甘肽还原酶(GR)酶活性提高, 过氧化氢(H2O2)、超氧阴离子自由基(O2?)和丙二醛(MDA)含量较低。与花前未进行渍水锻炼和花后渍水处理(NW)相比, 花前渍水锻炼和花后渍水处理(PW)下, 渍水锻炼敏感型品种较渍水锻炼不敏感品种显著提高了花后渍水胁迫下小麦旗叶SPAD (8.8%)和ΦPSII (17.6%)、降低非调节性能量耗散ΦNO (10.7%)和调节性能量耗散ΦNPQ (16.5%), 提升SOD (15.8%)、CAT (17.8%)、APX (8.9%)以及GR (30.7%)酶活性, 增加了叶片可溶性糖(17.5%)和蔗糖含量(21.6%), 促进花前贮藏物质向籽粒的转运率REP (20.0%), 同步提升PAA (10.8%)。与不耐渍型品种相比, 耐渍型小麦品种在花后渍水胁迫下旗叶的光合能力、抗氧化能力和干物质向籽粒的转运能力更强。花前渍水锻炼提高了各品种小麦花后渍水胁迫下旗叶的光合能力、抗氧化能力和干物质向籽粒的转运能力, 增强了小麦耐渍性; 与渍水锻炼不敏感型品种相比, 渍水锻炼敏感型品种的光合能力和抗氧化酶活性增幅较大。
| [1] | Singh G, Kumar P, Gupta V, Tyagi B S, Singh C, Kumar S A, Singh G P. Multivariate approach to identify and characterize bread wheat (Triticum aestivum L.) germplasm for waterlogging tolerance in India. Field Crops Res, 2018, 221:81-89. |
| [2] | Chen Y Y, Huang J F, Song X D, Gao P. Spatiotemporal characteristics of winter wheat waterlogging in the middle and lower reaches of the Yangtze River, China. Adv Meteorol, 2018, 9:1-11. |
| [3] | 任正隆. 中国南方小麦优质高效生产的若干问题. 四川农业大学学报, 2002, 20:299-303. |
| Ren Z L. Several limiting factors of wheat production in south area of China and the new approach of wheat breeding. J Sichuan Agric Univ, 2002, 20:299-303 (in Chinese with English abstract). | |
| [4] | Crisp P A, Ganguly D, Eichten S R. Reconsidering plant memory: intersections between stress recovery, RNA turnover, and epigenetics. Sci Adv, 2016, 2:e1501340. |
| [5] | Wang X, Liu F L, Jiang D. Priming: a promising strategy for crop production in response to future climate. J Integr Agric, 2017, 16:2709-2716. |
| [6] | 李同花, 王笑, 蔡剑, 周琴, 戴廷波, 姜东. 不同小麦品种对干旱锻炼响应的综合评价. 麦类作物学报, 2018, 38(1):65-73. |
| Li T H, Wang X, Cai J, Zhou Q, Dai T B, Jiang D. Comprehensive evaluation of drought priming on plant tolerance in different wheat cultivars. J Triticeae Crops, 2018, 38(1):65-73 (in Chinese with English abstract). | |
| [7] | Zhou W G, Chen F, Meng Y J, Chandrasekaran U, Shu K. Plant waterlogging/flooding stress responses: from seed germination to maturation. Plant Physiol Bioch, 2020, 148:228-236. |
| [8] | Herzog M, Striker G G, Colmer T D, Pedersen O. Mechanisms of waterlogging tolerance in wheat: a review of root and shoot physiology. Plant Cell Environ, 2016, 39:1068-1086. |
| [9] | Li C Y, Jiang D, Wollenweber B, Li Y, Dai T B, Cao W X. Waterlogging pretreatment during vegetative growth improves tolerance to waterlogging after anthesis in wheat. Plant Sci, 2011, 180:672-678. |
| [10] | Wang X, Huang M, Zhou Q, Cai J, Dai T B, Jiang D. Physiological and proteomic mechanisms of waterlogging priming improve tolerance to waterlogging stress in wheat (Triticum aestivum L.). Environ Exp Bot, 2016, 132:175-182. |
| [11] | Moya J L, Ros R, Picazo I. Influence of cadmium and nickel on growth, net photosynthesis and carbohydrate distribution in rice plants. Photosynth Res, 1993, 36:75-80. |
| [12] | 张永清, 苗果园, 李慧明. 冬小麦根系对不同生育时期渍水胁迫的生物学响应. 麦类作物学报, 2005, 25(4):59-63. |
| Zhang Y Q, Miao G Y, Li H M. Biological response of winter wheat root system to waterlogging stress at different growth stages. J Triticeae Crops, 2005, 25(4):59-63 (in Chinese with English abstract). | |
| [13] | Du Z Y, Bramlage W J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant-tissue extracts. J Agric Food Chem, 1992, 40:1566-1570. |
| [14] | Vwioko E, Adinkwu O, El-Esawi M A. Comparative physiological, biochemical, and genetic responses to prolonged waterlogging stress in okra and maize given exogenous ethylene priming. Front Physiol, 2017, 8:1-10. |
| [15] | Tan W, Liu J, Dai T B, Cao W X, Jiang D. Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post-anthesis waterlogging. Photosynthetica, 2008, 46:21-27. |
| [16] | Liu N, Lin Z F, Guan L L, Gaughan G, Lin G Z. Antioxidant enzymes regulate reactive oxygen species during pod elongation in Pisum sativum and Brassica chinensis. PLoS One, 2014: e875882. |
| [17] | Thomas M A. Measuring activities of the enzymes superoxide dismutase and glutathione reductase in lichens. In: Kranner I C, Beckett R P, Varma A K, eds. Protocols in Lichenology. Springer Lab Manuals. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-56359-1_12. |
| [18] | Foyer C H, Halliwell B. The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta, 1976, 133:21-25. |
| [19] | Mishra S, Agrawal S B. Interactive effects between supplemental ultraviolet-B radiation and heavy metals on the growth and biochemical characteristics of Spinacia oleracea L. Brazilian J Plant Physiol, 2006, 18:307-314. |
| [20] | 姜东, 陶勤南, 张国平. 渍水对小麦扬麦5号旗叶和根系衰老的影响. 应用生态学报, 2002, 13:1519-1521. |
| Jiang D, Tao Q N, Zhang G P. Effect of waterlogging on senescence of flag leaf and root of wheat Yangmai 5. J Appl Ecol, 2002, 13:1519-1521 (in Chinese with English abstract). | |
| [21] | 武文明, 陈洪俭, 李金才, 魏凤珍, 王世济, 周向红. 氮肥运筹对孕穗期受渍冬小麦旗叶叶绿素荧光与籽粒灌浆特性的影响. 作物学报, 2012, 38:1088-1096. |
| Wu W M, Chen H J, Li J C, Wei F Z, Wang S J, Zhou X H. Effects of nitrogen fertilization application regime on dry matter, nitrogen accumulation and transportation in summer maize under waterlogging at the seedling Stage. Acta Agron Sin, 2012, 38:1088-1096 (in Chinese with English abstract). | |
| [22] | 胡继超, 曹卫星, 姜东, 罗卫红. 小麦水分胁迫影响因子的定量研究: I. 干旱和渍水胁迫对光合、蒸腾及干物质积累与分配的影响. 作物学报, 2004, 30:315-320. |
| Hu J C, Cao W X, Jiang D, Luo W H. Quantification of water stress factor for crop growth simulation: I. Effects of drought and waterlogging stress on photosynthesis, transpiration and dry matter partitioning in winter wheat. Acta Agron Sin, 2004, 30:315-320 (in Chinese with English abstract). | |
| [23] | 姜东, 谢祝捷, 曹卫星, 戴廷波, 荆奇. 花后干旱和渍水对冬小麦光合特性和物质运转的影响. 作物学报, 2004, 30:175-182. |
| Jiang D, Xie Z J, Cao W X, Dai T B, Jing Q. Effects of post-anthesis drought and waterlogging on photosynthetic characteristics, assimilates transportation in winter wheat. Acta Agron Sin, 2004, 30:175-182 (in Chinese with English abstract). | |
| [24] | Wang X, Cai J, Liu F, Jin M, Yu H, Jiang D, Wollenweber B, Dai T B, Cao W X. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J Cereal Sci, 2012, 55:331-336. |
| [25] | Bansal R, Srivastava J P. Antioxidative responses to short term waterlogging stress in pigeon pea. Ind J Plant Physiol, 2015, 20:182-185. |
| [26] | Candan N, Tarhan L. Tolerance or sensitivity responses of mentha pulegium to osmotic and waterlogging stress in terms of antioxidant defense systems and membrane lipid peroxidation. Environ Exp Bot, 2012, 75:83-88. |
| [27] | Sairam R K, Kumutha D, Ezhilmathi K, Chinnusamy V, Meena R C. Waterlogging induced oxidative stress and antioxidant activity in pigeonpea genotypes. Biol Plant, 2009, 53:75-84. |
| [28] | Oukarroum A, Bussotti F, Goltsev V, Kalaji H M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ Exp Bot, 2015, 109:80-88. |
| [29] | Kafi M, Stewart W S, Borland A M. Carbohydrate and proline contents in leaves, roots, and apices of salt-tolerant and salt sensitive wheat varieties. Russ J Plant Physl, 2003, 50:155-162. |
| [30] | Hossain A, Uddin S N. Mechanisms of waterlogging tolerance in wheat: Morphological and metabolic adaptations under hypoxia or anoxia. Aust J Crop Sci, 2011, 5:1094-1101. |
| [31] | Ren B Z, Zhang J W, Dong S, Liu P, Zhao B. Responses of carbon metabolism and antioxidant system of summer maize to waterlogging at different stages. J Agron Crop Sci, 2018, 204:505-514. |
| [1] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [2] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [3] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [4] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
| [5] | 刘运景, 郑飞娜, 张秀, 初金鹏, 于海涛, 代兴龙, 贺明荣. 宽幅播种对强筋小麦籽粒产量、品质和氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 716-725. |
| [6] | 马红勃, 刘东涛, 冯国华, 王静, 朱雪成, 张会云, 刘静, 刘立伟, 易媛. 黄淮麦区Fhb1基因的育种应用[J]. 作物学报, 2022, 48(3): 747-758. |
| [7] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [8] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
| [9] | 徐龙龙, 殷文, 胡发龙, 范虹, 樊志龙, 赵财, 于爱忠, 柴强. 水氮减量对地膜玉米免耕轮作小麦主要光合生理参数的影响[J]. 作物学报, 2022, 48(2): 437-447. |
| [10] | 孟颖, 邢蕾蕾, 曹晓红, 郭光艳, 柴建芳, 秘彩莉. 小麦Ta4CL1基因的克隆及其在促进转基因拟南芥生长和木质素沉积中的功能[J]. 作物学报, 2022, 48(1): 63-75. |
| [11] | 韦一昊, 于美琴, 张晓娇, 王露露, 张志勇, 马新明, 李会强, 王小纯. 小麦谷氨酰胺合成酶基因可变剪接分析[J]. 作物学报, 2022, 48(1): 40-47. |
| [12] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [13] | 罗江陶, 郑建敏, 蒲宗君, 范超兰, 刘登才, 郝明. 四倍体小麦与六倍体小麦杂种的染色体遗传特性[J]. 作物学报, 2021, 47(8): 1427-1436. |
| [14] | 王艳朋, 凌磊, 张文睿, 王丹, 郭长虹. 小麦B-box基因家族全基因组鉴定与表达分析[J]. 作物学报, 2021, 47(8): 1437-1449. |
| [15] | 习玲, 王昱琦, 朱微, 王益, 陈国跃, 蒲宗君, 周永红, 康厚扬. 78份四川小麦育成品种(系)条锈病抗性鉴定与抗条锈病基因分子检测[J]. 作物学报, 2021, 47(7): 1309-1323. |
|
||