欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (4): 1027-1034.doi: 10.3724/SP.J.1006.2022.14066
• 研究简报 • 上一篇
刘嘉欣1,2( ), 兰玉1,2, 徐倩玉1, 李红叶1, 周新宇3, 赵璇1, 甘毅1, 刘宏波1, 郑月萍1, 詹仪花1, 张刚3, 郑志富1,2,*()
), 兰玉1,2, 徐倩玉1, 李红叶1, 周新宇3, 赵璇1, 甘毅1, 刘宏波1, 郑月萍1, 詹仪花1, 张刚3, 郑志富1,2,*()
LIU Jia-Xin1,2(), LAN Yu1,2, XU Qian-Yu1, LI Hong-Ye1, ZHOU Xin-Yu3, ZHAO Xuan1, GAN Yi1, LIU Hong-Bo1, ZHENG Yue-Ping1, ZHAN Yi-Hua1, ZHANG Gang3, ZHENG Zhi-Fu1,2,*()
摘要:
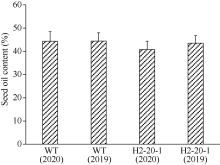
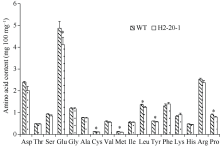
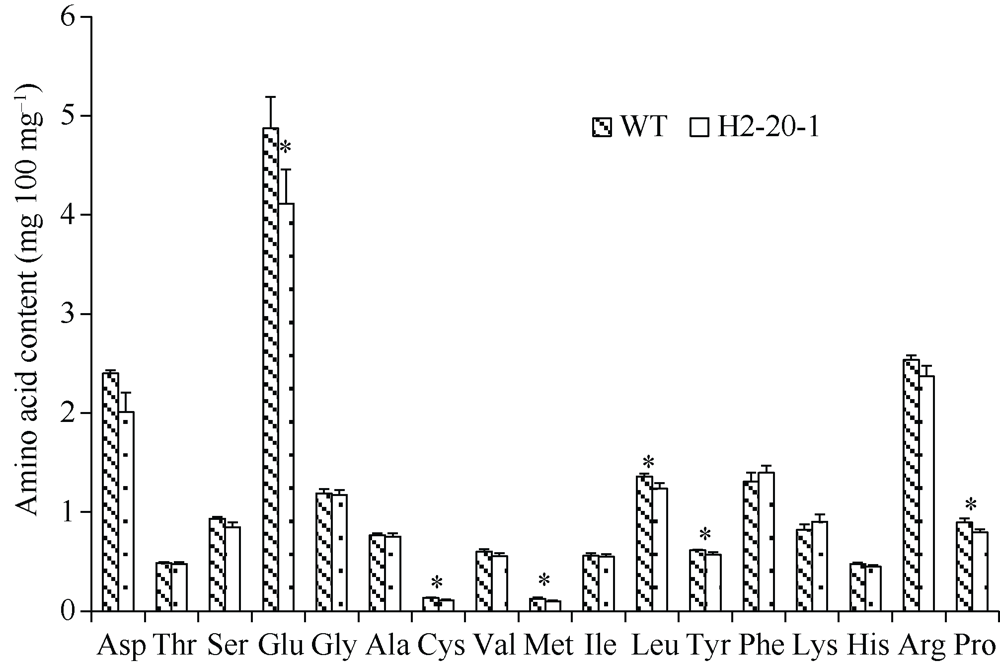
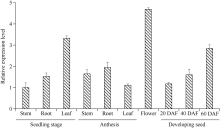
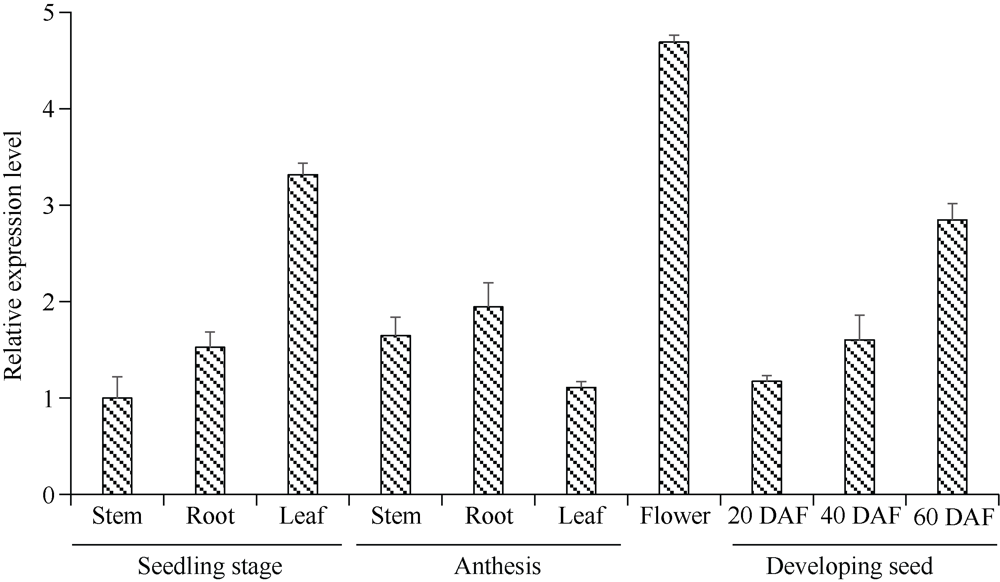
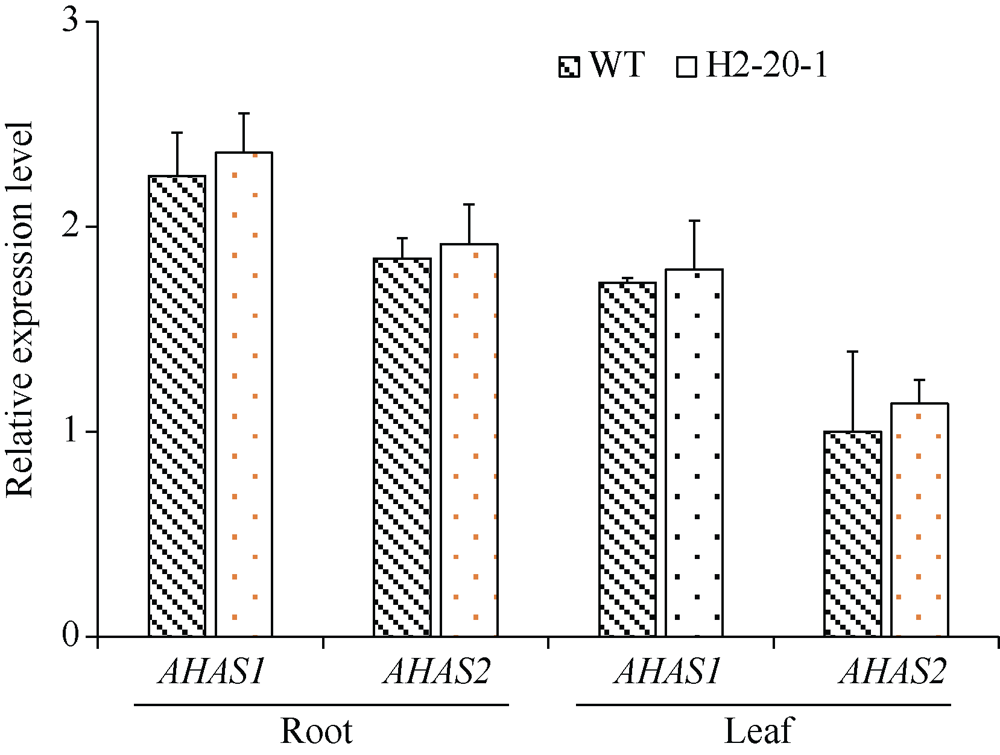
目前耐除草剂花生的种质资源匮乏, 这制约了花生栽培模式的多元化发展。为创制耐不同除草剂的花生种质资源, 我们利用甲基磺酸乙酯诱变技术创建了一个由55,000多个花生株系组成的诱变群体, 随后选择多种不同除草剂对该群体进行筛选, 获得多个对不同除草剂具有耐性的花生突变体。其中一个突变体在田间叶面喷施处理和多种实验室条件下的除草剂耐受性评估试验中均表现出对唑嘧磺草胺和双氟磺草胺极强的耐性, 但这种耐性性状并未对花生的产量与品质产生不良影响。为明确这种性状是否与除草剂靶标抗性相关联, 我们比较分析了该突变体与野生型中这2种除草剂靶标酶(乙酰羟酸合酶, AHAS)的基因序列及其表达量的差异。分子克隆与序列分析显示, 花生A10与B10染色体上各含一个与拟南芥AtAHAS序列高度相似的基因, 分别命名为AhAHAS1a与AhAHAS1b。花生A08与B08染色体上亦各含一个AHAS基因, 分别命名为AhAHAS2a与AhAHAS2b。然而, 与野生型相比, 突变体中的这些基因并未发生能够致使氨基酸序列发生改变的核苷酸替换。进而发现, AhAHAS基因的表达量在突变体与野生型中没有显著差异。这些结果表明, 该花生突变体的除草剂耐性可能由非靶标抗性机制所致。
| [1] | 万书波. 我国花生产业面临的机遇与科技发展战略. 中国农业科技导报, 2009, 11(1):7-12. |
| Wan S B. Opportunities facing peanut industry in China and strategies for its science and technology development. J Agric Sci Technol, 2009, 11(1):7-12 (in Chinese with English abstract). | |
| [2] | 丁小霞, 李培武, 周海燕, 李娟, 白艺珍. 花生农药最大残留限量标准比对研究. 中国油料作物学报, 2011, 33:527-531. |
| Ding X X, Li P W, Zhou H Y, Li J, Bai Y Z. Comparative study on maximum residue limits standards of pesticides in peanuts. Chin J Oil Crop Sci, 2011, 33:527-531 (in Chinese with English abstract). | |
| [3] |
Jabusch T W, Tjeerdema R S. Chemistry and fate of triazolopyrimidine sulfonamide herbicides. Rev Environ Contam Toxicol, 2008, 193:31-52.
doi: 10.1007/978-0-387-73163-6_2 pmid: 20614343 |
| [4] |
Cui H L, Li X J, Wang G Q, Wang J P, Wei S H, Cao H Y. Acetolactate synthase proline (197) mutations confer tribenuron-methyl resistance in Capsella bursa-pastoris populations from China. Pestic Biochem Physiol, 2012, 102:229-232.
doi: 10.1016/j.pestbp.2012.01.007 |
| [5] |
Lee H, Ullrich S E, Burke I C, Yenish J, Paulitz T C. Interactions between the root pathogen Rhizoctonia solani AG-8 and acetolactate-synthase-inhibiting herbicides in barley. Pest Manag Sci, 2012, 68:845-852.
doi: 10.1002/ps.v68.6 |
| [6] |
Liu W, Bi Y, Li L, Yuan G, Wang J. Molecular basis of resistance to tribenuron in water starwort (Myosoton aquaticum) populations from China. Weed Sci, 2013, 61:390-395.
doi: 10.1614/WS-D-12-00200.1 |
| [7] |
徐倩玉, 兰玉, 刘嘉欣, 周新宇, 张刚, 郑志富. 乙酰羟酸合酶抑制剂类除草剂的植物抗性机制. 作物学报, 2019, 45:1295-1302.
doi: 10.3724/SP.J.1006.2019.93003 |
| Xu Q Y, Lan Y, Liu J X, Zhou X Y, Zhang G, Zheng Z F. Mechanisms underlying plant resistance to the acetohydroxyacid synthase-inhibiting herbicides. Acta Agron Sin, 2019, 45:1295-1302 (in Chinese with English abstract). | |
| [8] |
Tan S, Evans R R, Dahmer M L, Singh B K, Shaner D L. Imidazolinone-tolerant crops: history, current status and future. Pest Manag Sci, 2005, 61:246-257.
doi: 10.1002/(ISSN)1526-4998 |
| [9] |
Pozniak C J, Birk I T, O’Donoughue L S, Ménard C, Hucl P J, Singh B K, Physiological and molecular characterization of mutation-derived imidazolinone resistance in spring wheat. Crop Sci, 2004, 44:1434-1443.
doi: 10.2135/cropsci2004.1434 |
| [10] |
Lee H, Rustgi S, Kumar N, Burke I, Yenish J P, Gill K S, von Wettstein D, Ullrich S E. Single nucleotide mutation in the barley acetohydroxy acid synthase (AHAS) gene confers resistance to imidazolinone herbicides. Proc Natl Acad Sci USA, 2011, 108:8909-8913.
doi: 10.1073/pnas.1105612108 |
| [11] | Rajasekaran K, Grula J W, Anderson D M. Selection and charac-terization of mutant cotton (Gossypium hirsutum L.) cell lines resistant to sulfonylurea and imidazolinone herbicides. Plant Sci, 1996, 199:115-124. |
| [12] |
Ghio C, Ramos M L, Altieri E, Bulos M, Sala C A. Molecular characterization of Als1, an acetohydroxyacid synthase mutation conferring resistance to sulfonylurea herbicides in soybean. Theor Appl Genet, 2013, 126:2957-2968.
doi: 10.1007/s00122-013-2185-7 |
| [13] |
Walter K L, Strachan S D, Ferry N M, Albert H H, Castle L A, Sebastian S A. Molecular and phenotypic characterization of Als1 and Als2 mutations conferring tolerance to acetolactate synthase herbicides in soybean. Pest Manag Sci, 2014, 70:1831-1839.
doi: 10.1002/ps.3725 pmid: 24425499 |
| [14] | 高建芹, 浦惠明, 戚存扣, 张洁夫, 龙卫华, 胡茂龙, 陈松, 陈新军, 陈锋, 顾慧. 抗咪唑啉酮油菜种质的发现与鉴定. 植物遗传资源学报, 2010, 11:369-373. |
| Gao J Q, Pu H M, Qi C K, Zhang J F, Long W H, Hu M L, Chen S, Chen X J, Chen F, Gu H. Identification of imidazoli-done- resistant oilseed rape mutant. J Plant Genet Resour, 2010, 11:369-373 (in Chinese with English abstract). | |
| [15] |
Kolkman J M, Slabaugh M B, Bruniard J M, Berry S, Bushman B S, Olungu C, Maes N, Abratti G, Zambelli A, Miller J F, Leon A, Knapp S J. Acetohydroxyacid synthase mutations conferring resistance to imidazolinone or sulfonylurea herbicides in sunflower. Theor Appl Genet, 2004, 109:1147-1159.
pmid: 15309298 |
| [16] |
Sala C A, Bulos M, Echarte M, Whitt S R, Ascenzi R. Molecular and biochemical characterization of an induced mutation conferring imidazolinone resistance in sunflower. Theor Appl Genet, 2008, 118:105-112.
doi: 10.1007/s00122-008-0880-6 |
| [17] |
Wright T R, Penner D. Cell selection and inheritance of imidazolinone resistance in sugar beet (Beta vulgaris). Theor Appl Genet, 1998, 96:612-620.
doi: 10.1007/s001220050779 |
| [18] | 周超, 张勇, 路兴涛, 马冲, 吴翠霞, 宋敏, 张田田, 孔繁华. 8种土壤处理除草剂对花生田杂草的防除效果及安全性评价. 农药, 2019, 58(3):226-229. |
| Zhou C, Zhang Y, Lu X T, Ma C, Wu C X, Song M, Zhang T T, Kong F H. The control effects and safety of eight soil treatment herbicides in peanut field. Agrochem, 2019, 58(3):226-229 (in Chinese with English abstract). | |
| [19] | 唐永常, 李永超, 耿锐. 麦套花生可持续发展技术存在问题及对策. 河南农业, 2014, (19):47. |
| Tang Y C, Li Y C, Geng R. Problems and countermeasures of sustainable development technology of wheat-covered peanut. Henan Agric, 2014, (19):47 (in Chinese). | |
| [20] |
Zheng Z, Xia Q, Dauk M, Selvaraj G, Zou J. Arabidopsis AtGPAT1, a member of the membrane-bound glycerol-3- phosphate acyltransferase gene family, is essential for tapetum differentiation and male fertility. Plant Cell, 2003, 15:1872-1887.
doi: 10.1105/tpc.012427 |
| [21] |
Gan Y, Song Y, Chen Y, Liu H, Yang D, Xu Q, Zheng Z. Transcriptome analysis reveals a composite molecular map linked to unique seed oil profile of Neocinnamomum caudatum(Nees) Merr. BMC Plant Biol, 2018, 18:303.
doi: 10.1186/s12870-018-1525-9 pmid: 30477425 |
| [22] | 赵青山, 付颖, 叶非. 三唑并嘧啶磺酰胺类除草剂的研究概况. 植物保护, 2011, 37(2):14-19. |
| Zhao Q S, Fu Y, Ye F. Study summary of triazolo [1,5-a] pyrimidine-2-sulfonanilide herbicides. Plant Prot, 2011, 37(2):14-19 (in Chinese with English abstract). | |
| [23] | 刘伟堂. 小麦田牛繁缕(Myosoton aquaticum L. Moench.)对苯磺隆的抗性研究. 山东农业大学博士学位论文,山东泰安, 2015. |
| Liu W T. Study on the Resistance to Tribenuron-methyl in Water Chickweed (Myosoton aquaticum L. Moench.) in Wheat Fields. PhD Dissertation of Shandong Agricultural University, Tai’an, Shandong,China, 2015 (in Chinese with English abstract). | |
| [24] |
Shaner D L, Anderson P C, Stidham M A. Imidazolinones: potent inhibitors of acetohydroxyacid synthase. Plant Physiol, 1984, 76:545-546.
doi: 10.1104/pp.76.2.545 pmid: 16663878 |
| [25] |
Petit C, Duhieu B, Boucansaud K, Delye C. Complex genetic control of non-target-site-based resistance to herbicides inhibiting acetyl-coenzyme A carboxylase and acetolactate-synthase in Alopecurus myosuroides Huds. Plant Sci, 2010, 178:501-509.
doi: 10.1016/j.plantsci.2010.03.007 |
| [26] |
Delye C, Pernin F, Scarabel L. Evolution and diversity of the mechanisms endowing resistance to herbicides inhibiting acetolactate-synthase (ALS) in corn poppy (Papaver rhoeas L.). Plant Sci, 2011, 180:333-342.
doi: 10.1016/j.plantsci.2010.10.007 |
| [27] |
Scarabel L, Pernin F, Délye C. Occurrence, genetic control and evolution of non-target-site based resistance to herbicides inhibiting acetolactate synthase (ALS) in the dicot weedPapaver rhoeas. Plant Sci, 2015, 238:158-169.
doi: 10.1016/j.plantsci.2015.06.008 pmid: 26259184 |
| [28] |
Yang Q, Deng W, Li X, Yu Q, Bai L, Zheng M. Target-site and non-target-site based resistance to the herbicide tribenuron- methyl in flixweed (Descurainia sophia L.). BMC Genomics, 2016, 17:551-563.
doi: 10.1186/s12864-016-2915-8 |
| [29] |
Mei Y, Si C, Liu M, Qiu L, Zheng M. Investigation of resistance levels and mechanisms to nicosulfuron conferred by non-target- site mechanisms in large crabgrass (Digitaria sanguinalis L.) from China. Pestic Biochem Physiol, 2017, 141:84-89.
doi: S0048-3575(16)30210-3 pmid: 28911745 |
| [30] |
Rey-Caballero J, Menéndez J, Osuna M D, Salas M, Torra J. Target-site and non-target-site resistance mechanisms to ALS inhibiting herbicides in Papaver rhoeas. Pestic Biochem Physiol, 2017, 138:57-65.
doi: S0048-3575(17)30082-2 pmid: 28456305 |
| [31] |
Zhao B C, Fu D N, Yu Y, Huang C T, Yan K C, Li P S, Shafi J, Zhu H, Wei S H, Ji M S. Non-target-site resistance to ALS-inhibiting herbicides in a Sagittaria trifolia L. population. Pestic Biochem Physiol, 2017, 140:79-84.
doi: 10.1016/j.pestbp.2017.06.008 |
| [32] |
Siminszky B, Corbin F T, Ward E R, Fleischmann T J, Dewey R E. Expression of a soybean cytochrome P450 monooxygenase cDNA in yeast and tobacco enhances the metabolism of phenylurea herbicides. Proc Natl Acad Sci USA, 1999, 96:1750-1755.
doi: 10.1073/pnas.96.4.1750 |
| [33] |
Saika H, Horita J, Taguchi-Shiobara F, Nonaka S, Nishizawa-Yokoi A, Iwakami S, Hori K, Matsumoto T, Tanaka T, Itoh T, Yano M, Kaku K, Shimizu T, Toki S. A novel rice cytochrome P450 gene,CYP72A31, confers tolerance to acetolactate synthase-inhibiting herbicides in rice and Arabidopsis. Plant Physiol, 2014, 166:1232-1240.
doi: 10.1104/pp.113.231266 |
| [1] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [2] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [3] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [4] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [5] | 汪颖, 高芳, 刘兆新, 赵继浩, 赖华江, 潘小怡, 毕晨, 李向东, 杨东清. 利用WGCNA鉴定花生主茎生长基因共表达模块[J]. 作物学报, 2021, 47(9): 1639-1653. |
| [6] | 王建国, 张佳蕾, 郭峰, 唐朝辉, 杨莎, 彭振英, 孟静静, 崔利, 李新国, 万书波. 钙与氮肥互作对花生干物质和氮素积累分配及产量的影响[J]. 作物学报, 2021, 47(9): 1666-1679. |
| [7] | 石磊, 苗利娟, 黄冰艳, 高伟, 张忠信, 齐飞艳, 刘娟, 董文召, 张新友. 花生AhFAD2-1基因启动子及5'-UTR内含子功能验证及其低温胁迫应答[J]. 作物学报, 2021, 47(9): 1703-1711. |
| [8] | 高芳, 刘兆新, 赵继浩, 汪颖, 潘小怡, 赖华江, 李向东, 杨东清. 北方主栽花生品种的源库特征及其分类[J]. 作物学报, 2021, 47(9): 1712-1723. |
| [9] | 张鹤, 蒋春姬, 殷冬梅, 董佳乐, 任婧瑶, 赵新华, 钟超, 王晓光, 于海秋. 花生耐冷综合评价体系构建及耐冷种质筛选[J]. 作物学报, 2021, 47(9): 1753-1767. |
| [10] | 薛晓梦, 吴洁, 王欣, 白冬梅, 胡美玲, 晏立英, 陈玉宁, 康彦平, 王志慧, 淮东欣, 雷永, 廖伯寿. 低温胁迫对普通和高油酸花生种子萌发的影响[J]. 作物学报, 2021, 47(9): 1768-1778. |
| [11] | 郝西, 崔亚男, 张俊, 刘娟, 臧秀旺, 高伟, 刘兵, 董文召, 汤丰收. 过氧化氢浸种对花生种子发芽及生理代谢的影响[J]. 作物学报, 2021, 47(9): 1834-1840. |
| [12] | 张旺, 冼俊霖, 孙超, 王春明, 石丽, 于为常. CRISPR/Cas9编辑花生FAD2基因研究[J]. 作物学报, 2021, 47(8): 1481-1490. |
| [13] | 戴良香, 徐扬, 张冠初, 史晓龙, 秦斐斐, 丁红, 张智猛. 花生根际土壤细菌群落多样性对盐胁迫的响应[J]. 作物学报, 2021, 47(8): 1581-1592. |
| [14] | 黄冰艳, 孙子淇, 刘华, 房元瑾, 石磊, 苗利娟, 张毛宁, 张忠信, 徐静, 张梦圆, 董文召, 张新友. 花生巢式群体的脂肪含量遗传分析[J]. 作物学报, 2021, 47(6): 1100-1108. |
| [15] | 许静, 潘丽娟, 李昊远, 王通, 陈娜, 陈明娜, 王冕, 禹山林, 侯艳华, 迟晓元. 花生油脂合成相关基因的表达谱分析[J]. 作物学报, 2021, 47(6): 1124-1137. |
|
||