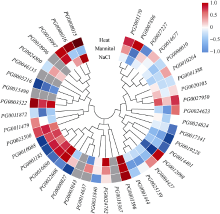
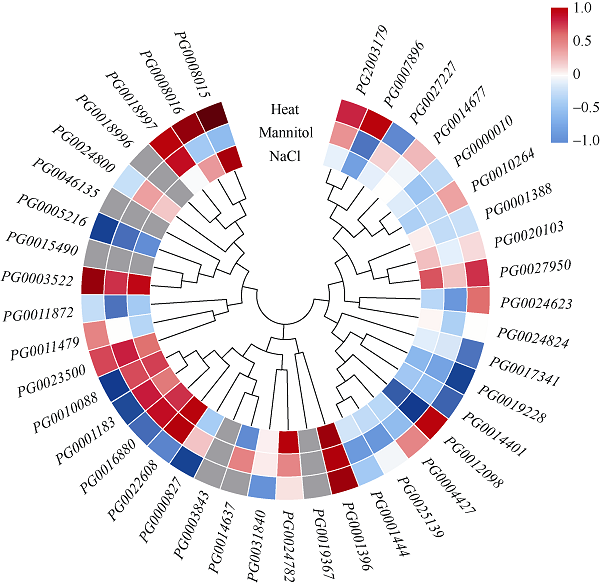
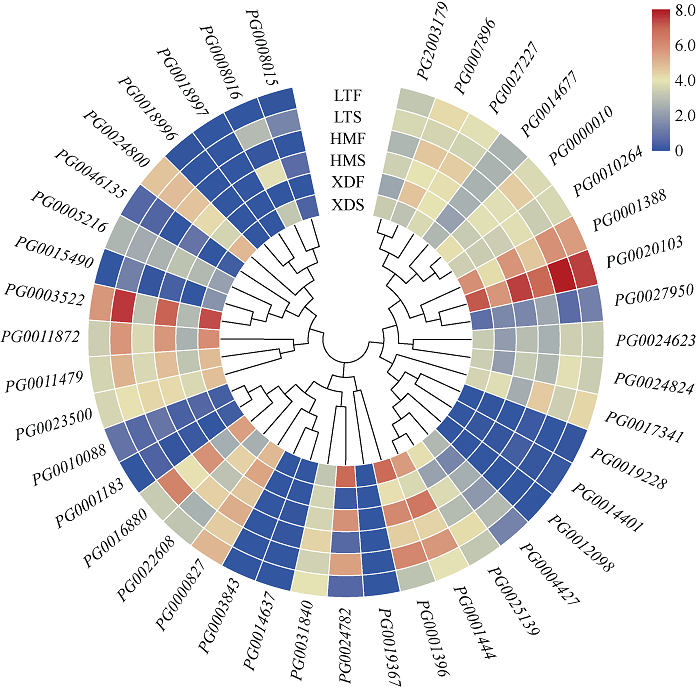
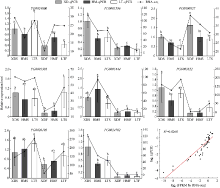
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2021, Vol. 47 ›› Issue (12): 2348-2361.doi: 10.3724/SP.J.1006.2021.04268
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
NIU Na1,2( ), LIU Zhen2, HUANG Peng-Xiang2,3, ZHU Jin-Yong1,2, LI Zhi-Tao1,2, MA Wen-Jing1,2, ZHANG Jun-Lian2,3, BAI Jiang-Ping1,2,*(), LIU Yu-Hui1,2,*()
), LIU Zhen2, HUANG Peng-Xiang2,3, ZHU Jin-Yong1,2, LI Zhi-Tao1,2, MA Wen-Jing1,2, ZHANG Jun-Lian2,3, BAI Jiang-Ping1,2,*(), LIU Yu-Hui1,2,*()
| [1] | 田鹏, 刘占林. 糖基转移酶超家族. 生命的化学, 2011, 31:732-736. |
| Tian P, Liu Z L. Glycosyltransferase superfamily. Chem Life, 2011, 31:732-736 (in Chinese with English abstract). | |
| [2] |
Vincent L, Hemalatha G R, Elodie D, Coutinho P M, Bernard H. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res, 2014, 42:D490-D495.
doi: 10.1093/nar/gkt1178 |
| [3] |
Yin Y, Chen H, Hahn M G, Mohnen D, Xu Y. Evolution and function of the plant cell wall synthesis-related glycosyltransferase family 8. Plant Physiol, 2010, 153:1729-1746.
doi: 10.1104/pp.110.154229 |
| [4] |
Cantarel B L, Coutinho P M, Corinne R, Thomas B, Vincent L, Bernard H. The carbohydrate-active enzymes database (CAZy): an expert resource for glycogenomics. Nucleic Acids Res, 2009, 37:D233-D238.
doi: 10.1093/nar/gkn663 |
| [5] |
Fabiana D G, Bermúdez L, Silvestre L B, Pereira D S A, Paula E, Demarco D, Alseekh S, Insani M, Buckeridge M, Almeida J, Grigioni G, Robert F A, Carrari F, Rossi M. Galacturonosyltransferase 4 silencing alters pectin composition and carbon partitioning in tomato. J Exp Bot, 2017, 64:2449-2466.
doi: 10.1093/jxb/ert106 |
| [6] |
Sterling J, Atmodjo M, Inwood S, Kolli V, Quigley H, Hahn M, Mohnen D. Functional identification of an Arabidopsis pectin biosynthetic homogalacturonan galacturonosyltransferase. Proc Natl Acad Sci USA, 2006, 103:5236-5241.
doi: 10.1073/pnas.0600120103 |
| [7] |
Mohnen D. Pectin structure and biosynthesis. Curr Opin Plant Biol, 2008, 11:266-277.
doi: 10.1016/j.pbi.2008.03.006 pmid: 18486536 |
| [8] |
Wang L, Wang W, Wang Y Q, Liu Y Y, Wang J X, Zhang X Q, Ye D, Chen L Q. Arabidopsis galacturonosyltransferase GAUT13 and GAUT14 have redundant functions in pollen tube growth. Mol Plant, 2013, 6:1131-1148.
doi: 10.1093/mp/sst084 pmid: 23709340 |
| [9] | 张松雨, 刘正文, 张艳, 杨君, 马峙英, 王省芬. 海岛棉GAUT基因家族的鉴定及其在棉纤维发育中的表达分析. 植物遗传资源学报, 2018, 19:722-730. |
| Zhang S Y, Liu Z W, Zhang Y, Yang J, Ma Z Y, Wang X F. Genome-wide identification of GAUT gene family in Gossypium barbadense L. and expression analysis in fiber developmental stages. J Plant Genet Resour, 2018, 19:722-730 (in Chinese with English abstract). | |
| [10] |
Altschul S F, Madden T L, Schäffer A A, Zhang J, Zhang Z, Miller W, Lipman D J. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res, 1997, 25:3389-3402.
pmid: 9254694 |
| [11] |
Bjellqvist B, Hughes G J, Pasquali C, Paquet N, Ravier F, Sanchez J C, Frutiger S, Hochstrasser D. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences. Electrophoresis, 1993, 14:1023-1031.
pmid: 8125050 |
| [12] | Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins M R, Appel R D, Bairoch A. Protein identification and analysis tools on the ExPASy server. Proteomics Protocol Handbook, 2005, 53:571-607. |
| [13] | 沈知临, 许磊, 陈文, 吴楠, 蔡永萍, 林毅, 高俊山. 亚洲棉和雷蒙德氏棉MATE基因家族生物信息学及其同源基因在陆地棉中的表达分析. 棉花学报, 2016, 28:215-226. |
| Shen Z L, Xu L, Chen W, Wu N, Cai Y P, Lin Y, Gao J S. Bioinformatic analysis of the multidrug and toxic compound extrusion gene family in Gossypium arboreum and Gossypium raimondii and expression of orthologs in Gossypium hirsutum. Cotton Sci, 2016, 28:215-226 (in Chinese with English abstract). | |
| [14] |
Bailey T L, Mikael B, Buske F A, Martin F, Grant C E, Luca C, Ren J, Li W W, Noble W S. MEME Suite: tools for motif discovery and searching. Nucleic Acids Res, 2009, 37:W202-W208.
doi: 10.1093/nar/gkp335 |
| [15] | 郭安源, 朱其慧, 陈新, 罗静初. GSDS: 基因结构显示系统. 遗传, 2007, 29:1023-1026. |
| Guo A Y, Zhu Q H, Chen X, Luo J C. GSDS: a gene structure display server. Hereditas (Beijing), 2007, 29:1023-1026 (in Chinese with English abstract). | |
| [16] |
Zheng L G, Andre C, Feng C C, Peter B, Wen H L. Extent of gene duplication in the genomes of drosophila, nematode, and yeast. Mol Biol Evol, 2020, 19:256-262.
doi: 10.1093/oxfordjournals.molbev.a004079 |
| [17] |
Yang S, Zhang X, Yue J X, Tian D, Chen J Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol Genet Genomics, 2008, 280:187-198.
doi: 10.1007/s00438-008-0355-0 |
| [18] |
Wang L, Guo K, Li Y, Tu Y, Peng L. Expression profiling and integrative analysis of the CESA/CSL superfamily in rice. BMC Plant Biol, 2010, 10:282.
doi: 10.1186/1471-2229-10-282 |
| [19] |
Wang Y, Tang H, Debarry J D, Tan X, Li J, Wang X, Lee T H, Jin H, Barry M, Guo H. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res, 2012, 40:e49.
doi: 10.1093/nar/gkr1293 |
| [20] |
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, Jones S J, Marra M A. Circos: an information aesthetic for comparative genomics. Genome Res, 2009, 19:1639-1645.
doi: 10.1101/gr.092759.109 pmid: 19541911 |
| [21] |
Wang D, Zhang Y, Zhang Z, Zhu J, Yu J. KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies. Genomics Proteomics Bioinf, 2010, 8:77-80.
doi: 10.1016/S1672-0229(10)60008-3 |
| [22] |
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol, 2016, 33:1870-1874.
doi: 10.1093/molbev/msw054 |
| [23] |
Tang X, Zhang N, Si H, Calderón-Urrea A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods, 2017, 13:85.
doi: 10.1186/s13007-017-0238-7 pmid: 29075311 |
| [24] | Chen C, Xia R, Chen H, He Y. TBtools, a toolkit for biologists integrating various biological HTS-data handling tools with a user-friendly interface. BioRxiv, 2018, 6:289660. |
| [25] |
Cannon S B, Mitra A, Baumgarten A, Young N D, May G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol, 2004, 4:10.
doi: 10.1186/1471-2229-4-10 |
| [26] |
Hanada K, Zou C, Lehti-Shiu D M, Shinozaki K, Shiu S H. Importance of lineage-specific expansion of plant tandem duplicates in the adaptive response to environmental stimuli. Plant Physiol, 2008, 148:993-1003.
doi: 10.1104/pp.108.122457 |
| [27] |
Jiang S Y, Ma Z, Ramachandran S. Evolutionary history and stress regulation of the lectin superfamily in higher plants. BMC Evol Biol, 2010, 10:79.
doi: 10.1186/1471-2148-10-79 |
| [28] |
Dias A P, Braun E L, McMullen M D, Grotewold E. Recently duplicated maize R2R3 Myb genes provide evidence for distinct mechanisms of evolutionary divergence after duplication. Plant Physiol, 2003, 131:610-620.
doi: 10.1104/pp.012047 |
| [29] |
Caffall K H, Mohnen D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydrate Res, 2009, 344:1879-1900.
doi: 10.1016/j.carres.2009.05.021 |
| [30] |
Xu G, Guo C, Shan H, Kong H. Divergence of duplicate genes in exon-intron structure. Proc Natl Acad Sci USA, 2012, 109:1187-1192.
doi: 10.1073/pnas.1109047109 |
| [31] |
Xiao J, Hu R, Gu T, Han J, Qiu D, Su P, Feng J, Chang J, Yang G, He G. Genome-wide identification and expression profiling of trihelix gene family under abiotic stresses in wheat. BMC Genomics, 2019, 20:287.
doi: 10.1186/s12864-019-5632-2 pmid: 30975075 |
| [32] | Hao Z, Avci U, Tan L, Zhu X, Glushka J, Pattathil S, Eberhard S, Sholes T, Rothstein G E, Lukowitz W. Loss of Arabidopsis GAUT12/IRX8 causes anther indehiscence and leads to reduced G lignin associated with altered matrix polysaccharide deposition. Front Plant Sci, 2014, 5:357. |
| [1] | XIE Qin-Qin, ZUO Tong-Hong, HU Deng-Ke, LIU Qian-Ying, ZHANG Yi-Zhong, ZHANG He-Cui, ZENG Wen-Yi, YUAN Chong-Mo, ZHU Li-Quan. Molecular cloning and expression analysis of BoPUB9 in self-incompatibility Brassica oleracea [J]. Acta Agronomica Sinica, 2022, 48(1): 108-120. |
|
||