欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (9): 2398-2411.doi: 10.3724/SP.J.1006.2023.22059
刘凯1( ), 陈积金1, 刘帅2, 陈旭1, 赵新茹2, 孙尚2, 薛超2, 龚志云1,2,*()
), 陈积金1, 刘帅2, 陈旭1, 赵新茹2, 孙尚2, 薛超2, 龚志云1,2,*()
LIU Kai1(), CHEN Ji-Jin1, LIU Shuai2, CHEN Xu1, ZHAO Xin-Ru2, SUN Shang2, XUE Chao2, GONG Zhi-Yun1,2,*()
摘要:
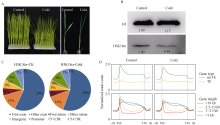
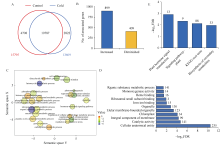
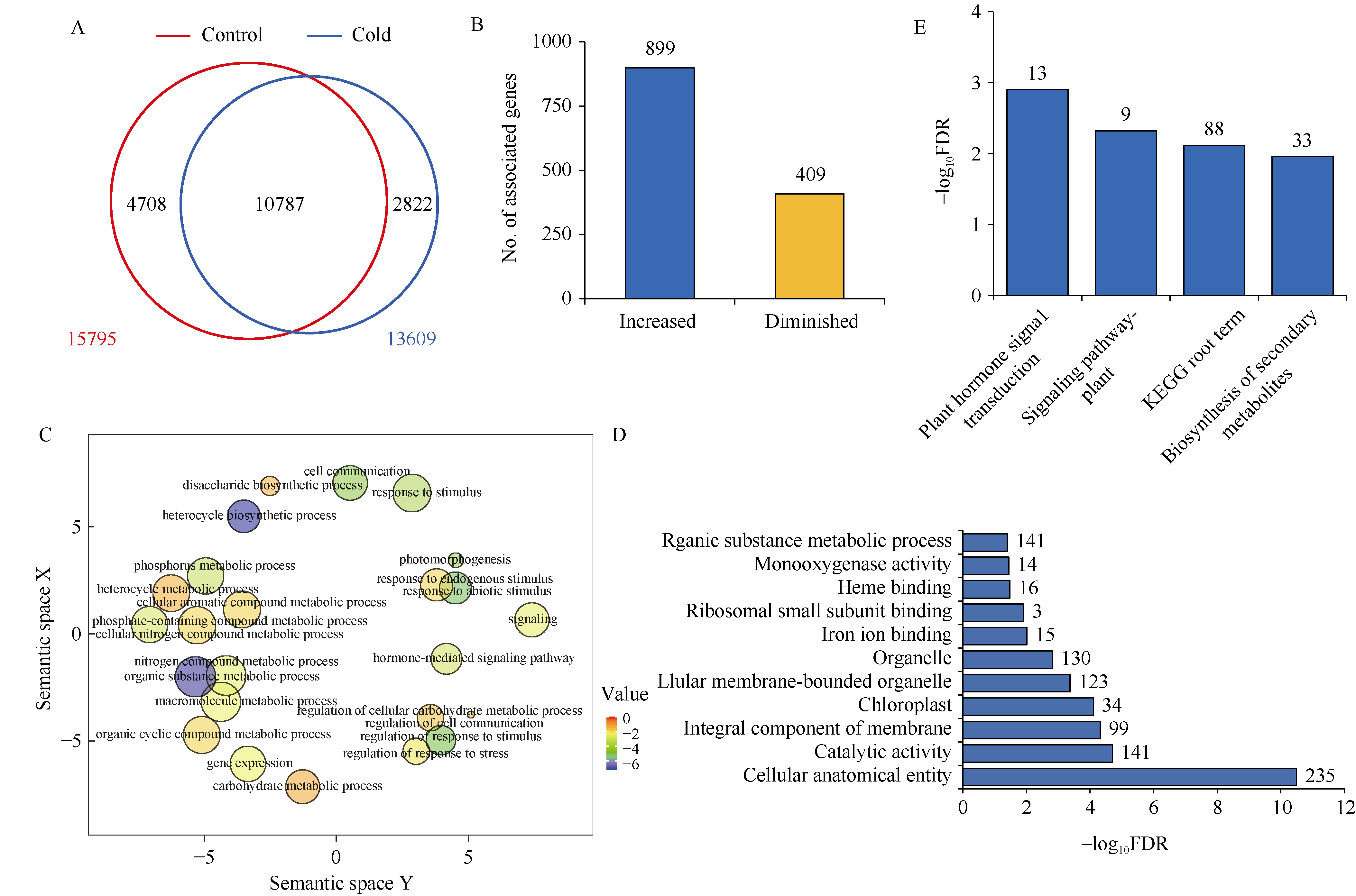
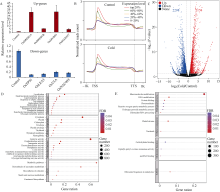
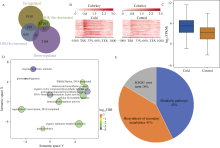
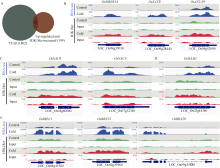
组蛋白修饰在水稻响应非生物胁迫的过程中发挥着重要作用。巴豆酰化是一种新型的组蛋白修饰方式, 其在水稻受到低温逆境时如何变化目前很少有见报道。本研究对正常生长和低温处理的水稻日品种本晴幼苗进行RNA-seq和ChIP-seq高通量测序, 然后联合分析组蛋白H3赖氨酸18特异位点上巴豆酰化修饰(H3K18cr)在低温胁迫下对基因表达的调控特征。研究表明, 在基因组中H3K18cr主要富集在第1外显子和基因间区, 且与基因表达和基因长度呈现正相关。低温胁迫下, H3K18cr在水稻基因组上的分布区域没有变化, 但是蛋白免疫印记和ChIP-seq结果均表明整体修饰水平下降; 差异修饰分析发现低温胁迫后有899个和409个基因分别表现出修饰显著增加和减少。通过与RNA-seq关联分析显示共有199个基因H3K18cr修饰水平增高且表达水平上调, GO富集分析发现这些基因主要参与转录活性的调控等过程。进一步验证表明组蛋白H3K18cr通过调控OsDREB1A、OsEATB、OsAP2-39、OsNAC9等转录因子的表达来参与水稻低温胁迫的响应过程。相关研究结果为解析组蛋白巴豆酰化调控植物响应低温胁迫的表观遗传机制提供理论基础。
| [1] |
Zhou Y, Simpson S, Holloway A F, Charlesworth J, van der Mei I, Taylor B V. The potential role of epigenetic modifications in the heritability of multiple sclerosis. Mult Scler, 2014, 20: 135-140.
doi: 10.1177/1352458514520911 |
| [2] |
Peach S E, Rudomin E L, Udeshi N D, Carr S A, Jaffe J D. Quantitative assessment of chromatin immunoprecipitation grade antibodies directed against histone modifications reveals patterns of co-occurring marks on histone protein molecules. Mol Cell Proteomics, 2012, 11: 128-137.
doi: 10.1074/mcp.M111.015941 pmid: 22442256 |
| [3] |
Kouzarides T. Chromatin modifications and their function. Cell, 2007, 128: 693-705.
doi: 10.1016/j.cell.2007.02.005 pmid: 17320507 |
| [4] | 薛超. 水稻盐胁迫下组蛋白乙酰化修饰特征及HATs相关基因的功能研究. 扬州大学博士学位论文, 江苏扬州, 2018. |
| Xue C. Characteristic Analysis of Histone Acetylation Dynamics under Salt Stress and Functional Analysis of HATs in Rice. PhD Dissertation of Yangzhou University, Yangzhou, Jiangsu, China, 2018. (in Chinese with English abstract) | |
| [5] |
Bao X C, Wang Y, Li X, Li X M, Liu Z, Yang T P, Wong C F, Zhang J W, Hao Q, Li X D. Identification of ‘erasers’ for lysine crotonylated histone marks using a chemical proteomics approach. eLife, 2014, 3: e02999.
doi: 10.7554/eLife.02999 |
| [6] | Tan M J, Luo H, Lee S, Jin F L, Yang J S, Montellier E, Buchou T, Cheng Z Y, Rousseaux S, Rajagopal N, Lu Z K, Ye Z, Zhu Q, Wysocka J, Ye Y, Khochbin S, Ren B, Zhao Y M. Identification of 67 histone marks and histone lysine crotonylation as a new type of histone modification. Cell, 2011, 146: 1015-1027. |
| [7] |
Sabari B R, Tang Z Y, Huang H, Yong-Gonzalez V, Molina H, Kong H E, Dai L Z, Shimada M, Cross J R, Zhao Y M, Roeder R G, Allis C D. Intracellular crotonyl-CoA stimulates transcription through p300-catalyzed histone crotonylation. Mol Cell, 2015, 58: 203-215.
doi: 10.1016/j.molcel.2015.02.029 pmid: 25818647 |
| [8] |
Sun H J, Liu X W, Li F F, Li W, Zhang J, Xiao Z X, Shen L L, Li Y, Wang F L, Yang J G. First comprehensive proteome analysis of lysine crotonylation in seedling leaves of Nicotiana tabacum. Sci Rep, 2017, 7: 3013.
doi: 10.1038/s41598-017-03369-6 |
| [9] |
Liu K D, Yuan C C, Li H L, Chen K Y, Lu L S, Shen C J, Zheng X L. A qualitative proteome-wide lysine crotonylation profiling of papaya (Carica papaya L.). Sci Rep, 2018, 8: 8230.
doi: 10.1038/s41598-018-26676-y |
| [10] |
Sun J H, Qiu C, Qian W J, Wang Y, Sun L T, Li Y S, Ding Z T. Ammonium triggered the response mechanism of lysine crotonylome in tea plants. BMC Genomics, 2019, 20: 340.
doi: 10.1186/s12864-019-5716-z pmid: 31060518 |
| [11] |
Dong Y B, Chen C. Comprehensive profiling of paper mulberry (Broussonetia papyrifera) crotonylome reveals the significance of lysine crotonylation in young leaves. Int J Mol Sci, 2022, 23: 1173.
doi: 10.3390/ijms23031173 |
| [12] | Xu M L, Luo J D, Li Y, Shen L L, Zhang X, Yu J, Guo Z Q, Wu J X, Chi Y C, Yang J G. First comprehensive proteomics analysis of lysine crotonylation in leaves of peanut (Arachis hypogaea L.). Proteomics, 2021, 21: e2000156. |
| [13] |
Liu S, Xue C, Fang Y, Chen G, Peng X J, Zhou Y, Chen C, Liu G Q, Gu M H, Wang K, Zhang W L, Wu Y F, Gong Z Y. Global involvement of lysine crotonylation in protein modification and transcription regulation in rice. Mol Cell Proteomics, 2018, 17: 1922-1936.
doi: 10.1074/mcp.RA118.000640 pmid: 30021883 |
| [14] |
Yuan L Y, Liu X C, Luo M, Yang S G, Wu K Q. Involvement of histone modifications in plant abiotic stress responses. J Integr Plant Biol, 2013, 55: 892-901.
doi: 10.1111/jipb.12060 |
| [15] |
Li H, Yan S H, Zhao L, Tan J J, Zhang Q, Gao F, Wang P, Hou H L, Li L J. Histone acetylation associated up-regulation of the cell wall related genes is involved in salt stress induced maize root swelling. BMC Plant Biol, 2014, 14: 105.
doi: 10.1186/1471-2229-14-105 pmid: 24758373 |
| [16] |
Kim J M, To T K, Ishida J, Morosawa T, Kawashima M, Matsui A, Toyoda T, Kimura H, Shinozaki K, Seki M. Alterations of lysine modifications on the histone H3 N-tail under drought stress conditions in Arabidopsis thaliana. Plant Cell Physiol, 2008, 49: 1580-1588.
doi: 10.1093/pcp/pcn133 |
| [17] |
Van D K, Ding Y, Malkaram S, Riethoven J J M, Liu R, Yang J Y, Laczko P, Chen H, Xia Y N, Ladunga I, Avramova Z, Fromm M. Dynamic changes in genome-wide histone H3 lysine 4 methylation patterns in response to dehydration stress in Arabidopsis thaliana. BMC Plant Biol, 2010, 10: 238.
doi: 10.1186/1471-2229-10-238 |
| [18] |
Vlachonasios K E, Thomashow M F, Triezenberg S J. Disruption mutations of ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis growth, development, and gene expression. Plant Cell, 2003, 15: 626-638.
doi: 10.1105/tpc.007922 |
| [19] |
Kwon C S, Lee D, Choi G, Chung W I. Histone occupancy- dependent and -independent removal of H3K27 trimethylation at cold-responsive genes in Arabidopsis. Plant J, 2009, 60: 112-121.
doi: 10.1111/tpj.2009.60.issue-1 |
| [20] | Roy D, Paul A, Roy A, Ghosh R, Ganguly P, Chaudhuri S. Differential acetylation of histone H3 at the regulatory region of OsDREB1b promoter facilitates chromatin remodelling and transcription activation during cold stress. PLoS One, 2014, 9: e100343. |
| [21] |
Zheng D Y, Wang L, Chen L F, Pan X C, Lin K D, Fang Y, Wang X E, Zhang W L. Salt-responsive genes are differentially regulated at the chromatin levels between seedlings and roots in rice. Plant Cell Physiol, 2019, 60: 1790-1803.
doi: 10.1093/pcp/pcz095 pmid: 31111914 |
| [22] |
Lu Y, Xu Q T, Liu Y, Yu Y, Cheng Z Y, Zhao Y, Zhou D X. Dynamics and functional interplay of histone lysine butyrylation, crotonylation, and acetylation in rice under starvation and submergence. Genome Biol, 2018, 19: 144.
doi: 10.1186/s13059-018-1533-y pmid: 30253806 |
| [23] |
徐青山, 黄晶, 孙爱军, 洪小智, 朱练峰, 曹小闯, 孔亚丽, 金千瑜, 朱春权, 张均华. 低温影响水稻发育机理及调控途径研究进展. 中国水稻科学, 2022, 36: 118-130.
doi: 10.16819/j.1001-7216.2022.210602 |
|
Xu Q S, Huang J, Sun A J, Hong X Z, Zhu L F, Cao X C, Kong Y L, Jin Q Y, Quan C, Zhang J H. Effects of low temperature on the growth and development of rice plants and the advance of regulation pathways: a review. Chin J Rice Sci, 2022, 36: 118-130. (in Chinese with English abstract)
doi: 10.16819/j.1001-7216.2022.210602 |
|
| [24] |
Hu Y F, Zhu N, Wang X M, Yi Q P, Zhu D Y, Lai Y, Zhao Y. Analysis of rice Snf2 family proteins and their potential roles in epigenetic regulation. Plant Physiol Biochem, 2013, 70: 33-42.
doi: 10.1016/j.plaphy.2013.05.001 |
| [25] |
Sharma R, Singh R K M, Malik G, Deveshwar P, Tyagi A K, Kapoor S, Kapoor M. Rice cytosine DNA methyltransferases: gene expression profiling during reproductive development and abiotic stress. FEBS J, 2009, 276: 6301-6311.
doi: 10.1111/j.1742-4658.2009.07338.x pmid: 19788421 |
| [26] |
Xue C, Liu S, Chen C, Zhu J, Yang X B, Zhou Y, Guo R, Liu X Y, Gong Z Y. Global proteome analysis links lysine acetylation to diverse functions in Oryza sativa. Proteomics, 2018, 18: 1700036.
doi: 10.1002/pmic.v18.1 |
| [27] |
Nagaki K, Talbert P B, Zhong C X, Dawe R K, Henikoff S, Jiang J M. Chromatin immunoprecipitation reveals that the 180-bp satellite repeat is the key functional DNA element of Arabidopsis thaliana centromeres. Genetics, 2003, 163: 1221-1225.
doi: 10.1093/genetics/163.3.1221 |
| [28] |
Ito Y, Katsura K, Maruyama K, Taji T, Kobayashi M, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol, 2006, 47: 141-153.
doi: 10.1093/pcp/pci230 pmid: 16284406 |
| [29] |
Mao D H, Chen C Y. Colinearity and similar expression pattern of rice DREB1s reveal their functional conservation in the cold-responsive pathway. PLoS One, 2012, 7: e47275.
doi: 10.1371/journal.pone.0047275 |
| [30] |
Huang L, Hong Y B, Zhang H J, Li D Y, Song F M. Rice NAC transcription factor ONAC095 plays opposite roles in drought and cold stress tolerance. BMC Plant Biol, 2016, 16: 203.
doi: 10.1186/s12870-016-0897-y pmid: 27646344 |
| [31] |
Liu H J, Li Q Z, Yang F, Zhu F Y, Sun Y, Tao Y Z, Lo C. Differential regulation of protochlorophyllide oxidoreductase abundances by VIRESCENT 5A (OsV5A) and VIRESCENT 5B (OsV5B) in rice seedlings. Plant Cell Physiol, 2016, 57: 2392-2402.
pmid: 27565208 |
| [32] |
He S, Tan L L, Hu Z L, Chen G P, Wang G X, Hu T Z. Molecular characterization and functional analysis by heterologous expression in E. coli under diverse abiotic stresses for OsLEA5, the atypical hydrophobic LEA protein from Oryza sativa L. Mol Genet Genomics, 2012, 287: 39-54.
doi: 10.1007/s00438-011-0660-x |
| [33] |
Li H W, Zang B S, Deng X W, Wang X P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta, 2011, 234: 1007-1018.
doi: 10.1007/s00425-011-1458-0 |
| [34] |
Yu C Y, Su S, Xu Y C, Zhao Y Q, Yan A, Huang L L, Ali I, Gan Y B. The effects of fluctuations in the nutrient supply on the expression of five members of the AGL17 clade of MADS-Box genes in rice. PLoS One, 2014, 9: e105597.
doi: 10.1371/journal.pone.0105597 |
| [35] |
Su C F, Wang Y C, Hsieh T H, Lu C A, Tseng T H, Yu S M. A novel MYBS3-dependent pathway confers cold tolerance in rice. Plant Physiol, 2010, 153: 145-158.
doi: 10.1104/pp.110.153015 |
| [36] |
Jin J P, Tian F, Yang D C, Meng Y Q, Kong L, Luo J C, Gao G. PlantTFDB 4.0: toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res, 2017, 45: D1040-D1045.
doi: 10.1093/nar/gkw982 |
| [37] |
Tripathi A K, Pareek A, Sopory S K, Singla-Pareek S L. Narrowing down the targets for yield improvement in rice under normal and abiotic stress conditions via expression profiling of yield- related genes. Rice, 2012, 5: 37.
doi: 10.1186/1939-8433-5-37 pmid: 24280046 |
| [38] |
Qi W W, Sun F, Wang Q J, Chen M L, Huang Y Q, Feng Y Q, Luo X J, Yang J S. Rice ethylene-response AP2/ERF factor OsEATB restricts internode elongation by down-regulating a gibberellin biosynthetic gene. Plant Physiol, 2011, 157: 216-228.
doi: 10.1104/pp.111.179945 |
| [39] |
Wan L Y, Zhang J F, Zhang H W, Zhang Z J, Quan R D, Zhou S R, Huang R F. Transcriptional activation of OsDERF1 in OsERF3 and OsAP2-39 negatively modulates ethylene synthesis and drought tolerance in rice. PLoS One, 2011, 6: e25216.
doi: 10.1371/journal.pone.0025216 |
| [40] |
Zhang X, Long Y, Chen X X, Zhang B L, Xin Y F, Li L Y, Cao S L, Liu F H, Wang Z G, Huang H, Zhou D G, Xia J X. A NAC transcription factor OsNAC3 positively regulates ABA response and salt tolerance in rice. BMC Plant Biol, 2021, 21: 546.
doi: 10.1186/s12870-021-03333-7 pmid: 34800972 |
| [41] |
Redillas M C F R, Jeong J S, Kim Y S, Jung H, Bang S W, Choi Y D, Ha S H, Reuzeau C, Kim J K. The overexpression of OsNAC9 alters the root architecture of rice plants enhancing drought resistance and grain yield under field conditions. Plant Biotechnol J, 2012, 10: 792-805.
doi: 10.1111/j.1467-7652.2012.00697.x pmid: 22551450 |
| [42] | Um T, Choi J, Park T, Chung P J, Jung S E, Shim J S, Kim Y S, Choi I Y, Park S C, Oh S J, Seo J S, Kim J K. Rice microRNA171f/SCL6 module enhances drought tolerance by regulation of flavonoid biosynthesis genes. Plant Direct, 2022, 6: e374. |
| [43] | 刘兰兰. 水稻OsBBX基因响应热胁迫的初步研究. 湖南农业大学硕士学位论文, 湖南长沙, 2015. |
| Liu L L. Preliminary Study on OsBBX Genes under Heat Stress in Rice. MS Thesis of Hunan Agricultural University, Changsha, Hunan, China, 2015. (in Chinese with English abstract) | |
| [44] |
刘焱, 邢立静, 李俊华, 戴绍军. 水稻含有B-box锌指结构域的OsBBX25蛋白参与植物对非生物胁迫的响应. 植物学报, 2012, 47: 366-378.
doi: 10.3724/SP.J.1259.2012.00366 |
| Liu Y, Xing L J, Li J H, Dai S J. Rice B-box zinc finger protein OsBBX25 is involved in the abiotic response. Chin Bull Bot, 2012, 47: 366-378. (in Chinese with English abstract) | |
| [45] |
Iwamoto M, Higo K, Takano M. Circadian clock- and phytochrome-regulated Dof-like gene, Rdd1, is associated with grain size in rice. Plant Cell Environ, 2009, 32: 592-603.
doi: 10.1111/pce.2009.32.issue-5 |
| [46] |
Gandass N, Kajal, Salvi P. Intrinsically disordered protein, DNA binding with one finger transcription factor (OsDOF27) implicates thermotolerance in yeast and rice. Front Plant Sci, 2022, 13: 956299.
doi: 10.3389/fpls.2022.956299 |
| [47] |
Kim S K, Yun C H, Lee J H, Jang Y H, Park H Y, Kim J K. OsCO3, a CONSTANS-LIKE gene, controls flowering by negatively regulating the expression of FT-like genes under SD conditions in rice. Planta, 2008, 228: 355-365.
doi: 10.1007/s00425-008-0742-0 |
| [48] | Lee Y S, Jeong D H, Lee D Y, Yi J, Ryu C H, Kim S L, Jeong H J, Choi S C, Jin P, Yang J, Cho L H, Choi H, An G. OsCOL4 is a constitutive flowering repressor upstream of Ehd1 and downstream of OsphyB. Plant J, 2010, 63: 18-30. |
| [49] |
Zhang S X, Haider I, Kohlen W, Jiang L, Bouwmeester H, Meijer A H, Schluepmann H, Liu C M, Ouwerkerk P B F. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol Biol, 2012, 80: 571-585.
doi: 10.1007/s11103-012-9967-1 |
| [50] |
Sun L, Di D W, Li G, Kronzucker H J, Wu X, Shi W. Endogenous ABA alleviates rice ammonium toxicity by reducing ROS and free ammonium via regulation of the SAPK9-bZIP20 pathway. J Exp Bot, 2020, 71: 4562-4577.
doi: 10.1093/jxb/eraa076 pmid: 32064504 |
| [51] |
Wang B X, Xu B, Liu Y, Li J F, Sun Z G, Chi M, Xing Y G, Yang B, Li J, Liu J B, Chen T M, Fang Z W, Lu B G, Xu D Y, Bello B K. A novel mechanisms of the signaling cascade associated with the SAPK10-bZIP20-NHX1 synergistic interaction to enhance tolerance of plant to abiotic stress in rice (Oryza sativa L.). Plant Sci, 2022, 323: 111393.
doi: 10.1016/j.plantsci.2022.111393 |
| [52] |
Mittal D, Chakrabarti S, Sarkar A, Singh A, Grover A. Heat shock factor gene family in rice: Genomic organization and transcript expression profiling in response to high temperature, low temperature and oxidative stresses. Plant Physiol Biochem, 2009, 47: 785-795.
doi: 10.1016/j.plaphy.2009.05.003 |
| [53] |
Ogawa S, Miyamoto K, Nemoto K, Sawasaki T, Yamane H, Nojiri H, Okada K. OsMYC2, an essential factor for JA-inductive sakuranetin production in rice, interacts with MYC2-like proteins that enhance its transactivation ability. Sci Rep, 2017, 7: 40175.
doi: 10.1038/srep40175 pmid: 28067270 |
| [54] | 李梦琪. 转录因子OsbHLH111抑制水稻幼苗生长机理的初步研究. 河南农业大学硕士学位论文, 河南郑州, 2018. |
| Li M Q. Preminary Study on the Mechanism for Rice Seedling Growth-inhibition of bHLH111. MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2018. (in Chinese with English abstract) | |
| [55] |
Tian Q X, Luan J X, Guo C X, Shi X Y, Deng P, Zhou Z Z, Zhang W H, Shen L K. A bHLH protein, OsBIM1, positively regulates rice leaf angle by promoting brassinosteroid signaling. Biochem Biophys Res Commun, 2021, 578: 129-135.
doi: 10.1016/j.bbrc.2021.09.035 |
| [56] |
Yin X, Cui Y, Wang M, Xia X. Overexpression of a novel MYB-related transcription factor, OsMYBR1, confers improved drought tolerance and decreased ABA sensitivity in rice. Biochem Biophys Res Commun, 2017, 490: 1355-1361.
doi: 10.1016/j.bbrc.2017.07.029 |
| [57] | Rongjun C. Isolation of a novel MYB transcription factor OsMyb1R from rice and analysis of the response of this gene to abiotic stresses. Afr J Biotechnol, 2012, 11: 3731-3737. |
| [58] |
Jha U C, Bohra A, Jha R, Parida S K. Salinity stress response and ‘omics’ approaches for improving salinity stress tolerance in major grain legumes. Plant Cell Rep, 2019, 38: 255-277.
doi: 10.1007/s00299-019-02374-5 |
| [59] |
Zeng Z X, Zhang W L, Marand A P, Zhu B, Buell C R, Jiang J M. Cold stress induces enhanced chromatin accessibility and bivalent histone modifications H3K4me3 and H3K27me3 of active genes in potato. Genome Biol, 2019, 20: 123.
doi: 10.1186/s13059-019-1731-2 pmid: 31208436 |
| [60] |
Sun L, Song G S, Guo W J, Wang W X, Zhao H K, Gao T T, Lyu Q X, Yang X, Xu F, Dong Y S, Pu L. Dynamic changes in genome-wide histone3 lysine27 trimethylation and gene expression of soybean roots in response to salt stress. Front Plant Sci, 2019, 10: 1031.
doi: 10.3389/fpls.2019.01031 pmid: 31552061 |
| [61] |
Lee H G, Seo P J. MYB96 recruits the HDA15 protein to suppress negative regulators of ABA signaling in Arabidopsis. Nat Commun, 2019, 10: 1713.
doi: 10.1038/s41467-019-09417-1 |
| [62] |
Ding Y, Avramova Z, Fromm M. The Arabidopsis trithorax-like factor ATX1 functions in dehydration stress responses via ABA-dependent and ABA-independent pathways. Plant J, 2011, 66: 735-744.
doi: 10.1111/j.1365-313X.2011.04534.x |
| [63] |
Pien S, Fleury D, Mylne J S, Crevillen P, Inze D, Avramova Z, Dean C, Grossniklaus U. ARABIDOPSIS TRITHORAX1 dynamically regulates FLOWERING LOCUS C activation via histone 3 lysine 4 trimethylation. Plant Cell, 2008, 20: 580-588.
doi: 10.1105/tpc.108.058172 |
| [64] |
Ullah F, Xu Q, Zhao Y, Zhou D X. Histone deacetylase HDA710 controls salt tolerance by regulating ABA signaling in rice. J Integr Plant Biol, 2021, 63: 451-467.
doi: 10.1111/jipb.13042 |
| [65] |
Nakashima K, Ito Y, Yamaguchi-Shinozaki K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol, 2009, 149: 88-95.
doi: 10.1104/pp.108.129791 pmid: 19126699 |
| [66] |
陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展. 作物学报, 2022, 48: 781-790.
doi: 10.3724/SP.J.1006.2022.12026 |
|
Chen Y, Sun M Z, Jia B W, Leng Y, Sun X L. Research progress regarding the function and mechanism of rice AP2/ERF transcription factor in stress response. Acta Agron Sin, 2022, 48: 781-790. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2022.12026 |
|
| [67] |
Zheng Y, Ge J, Bao C, Chang W, Liu J, Shao J, Liu X, Su L, Pan L, Zhou D X. Histone deacetylase HDA9 and WRKY53 transcription factor are mutual antagonists in regulation of plant stress response. Mol Plant, 2020, 13: 598-611.
doi: S1674-2052(19)30408-3 pmid: 31891777 |
| [68] |
Lamke J, Brzezinka K, Altmann S, Baurle I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J, 2016, 35: 162-175.
doi: 10.15252/embj.201592593 pmid: 26657708 |
| [69] | Mao Y, Pavangadkar K A, Thomashow M F, Triezenberg S J. Physical and functional interactions of Arabidopsis ADA 2 transcriptional coactivator proteins with the acetyltransferase GCN5 and with the cold-induced transcription factor CBF1. Biochim Biophys Acta, 2006, 1759: 69-79. |
| [70] |
Zong W, Yang J, Fu J, Xiong L. Synergistic regulation of drought-responsive genes by transcription factor OsbZIP23 and histone modification in rice. J Integr Plant Biol, 2020, 62: 723-729.
doi: 10.1111/jipb.12850 |
| [71] |
Huang Q X, Liao X Q, Yang X H, Luo Y C, Lin P, Zeng Q H, Bai H R, Jiang B B, Pan Y Z, Zhang F, Zhang L, Jia Y, Liu Q L. Lysine crotonylation of DgTIL1 at K72 modulates cold tolerance by enhancing DgnsLTP stability in chrysanthemum. Plant Biotechnol J, 2021, 19: 1125-1140.
doi: 10.1111/pbi.13533 pmid: 33368971 |
| [1] | 徐高峰, 申时才, 张付斗, 杨韶松, 金桂梅, 郑凤萍, 温丽娜, 张云, 吴冉迪. 土壤微生物对长雄野生稻及其化感潜力后代抑草作用的影响[J]. 作物学报, 2023, 49(9): 2562-2571. |
| [2] | 胡艳娟, 薛丹, 耿嫡, 朱末, 王天穹, 王晓雪. 水稻OsCDF1基因突变效应及其基因组变异分析[J]. 作物学报, 2023, 49(9): 2362-2372. |
| [3] | 唐杰, 龙湍, 吴春瑜, 李新鹏, 曾翔, 吴永忠, 黄培劲. 水稻OsGMS2基因的鉴定及其核不育系种子繁殖体系构建[J]. 作物学报, 2023, 49(8): 2025-2038. |
| [4] | 宋兆建, 冯紫旖, 屈天歌, 吕品苍, 杨晓璐, 湛明月, 张献华, 何玉池, 刘育华, 蔡得田. 四倍体水稻回复二倍体品系的籼粳属性鉴定和杂种优势利用初探[J]. 作物学报, 2023, 49(8): 2039-2050. |
| [5] | 韦新宇, 曾跃辉, 杨旺兴, 肖长春, 候新坡, 黄建鸿, 邹文广, 许旭明. 利用CRISPR-Cas9技术编辑Badh2基因创制优质香型籼稻三系不育系[J]. 作物学报, 2023, 49(8): 2144-2159. |
| [6] | 贾璐绮, 孙悠, 田然, 张学菲, 代永东, 崔志波, 李杨羊, 冯新宇, 桑贤春, 王晓雯. 水稻种子快速萌发突变体rgs1的鉴定及调控基因克隆[J]. 作物学报, 2023, 49(8): 2288-2295. |
| [7] | 丁洪艳, 冯晓溪, 汪柏宇, 张积森. 甘蔗割手密种LRRII-RLK基因家族演化和表达分析[J]. 作物学报, 2023, 49(7): 1769-1784. |
| [8] | 项嘉铭, 戴茜, 刘立军. 外源水杨酸提高云麻1号(Cannabis sativa L.)对铜胁迫的耐受性[J]. 作物学报, 2023, 49(7): 1979-1993. |
| [9] | 邓艾兴, 李歌星, 吕玉平, 刘猷红, 孟英, 张俊, 张卫建. 齐穗后遮阴时长对西北稻区粳稻产量和品质的影响[J]. 作物学报, 2023, 49(7): 1930-1941. |
| [10] | 许娜, 徐铨, 徐正进, 陈温福. 水稻株型生理生态与遗传基础研究进展[J]. 作物学报, 2023, 49(7): 1735-1746. |
| [11] | 魏正欣, 刘昌燕, 陈宏伟, 李莉, 孙龙清, 韩雪松, 焦春海, 沙爱华. 基于干旱胁迫转录组信息的蚕豆ASPAT基因家族分析[J]. 作物学报, 2023, 49(7): 1871-1881. |
| [12] | 林孝欣, 黄明江, 韦祎, 朱洪慧, 王子怡, 李忠成, 庄慧, 李彦羲, 李云峰, 陈锐. 水稻籽粒伸长突变体lgdp的鉴定与基因定位[J]. 作物学报, 2023, 49(6): 1699-1707. |
| [13] | 丁杰荣, 马雅美, 潘发枝, 江立群, 黄文洁, 孙炳蕊, 张静, 吕树伟, 毛兴学, 于航, 李晨, 刘清. 泛素受体蛋白OsDSK2b负向调控水稻叶瘟和渗透胁迫抗性[J]. 作物学报, 2023, 49(6): 1466-1479. |
| [14] | 何永明, 张芳. 生长素调控水稻颖花开放的效应研究[J]. 作物学报, 2023, 49(6): 1690-1698. |
| [15] | 陶玥玥, 盛雪雯, 徐坚, 沈园, 王海候, 陆长婴, 沈明星. 长三角水稻-油菜周年两熟温光资源分配与利用特征[J]. 作物学报, 2023, 49(5): 1327-1338. |
|