欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (2): 403-413.doi: 10.3724/SP.J.1006.2024.31016
范子培1,2( ), 李龙2, 史雨刚1, 孙黛珍1,*(), 李超男2,*(), 景蕊莲2
), 李龙2, 史雨刚1, 孙黛珍1,*(), 李超男2,*(), 景蕊莲2
FAN Zi-Pei1,2(), LI Long2, SHI Yu-Gang1, SUN Dai-Zhen1,*(), LI Chao-Nan2,*(), JING Rui-Lian2
摘要:
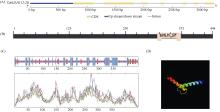
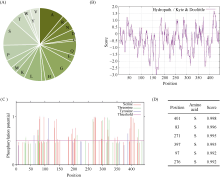
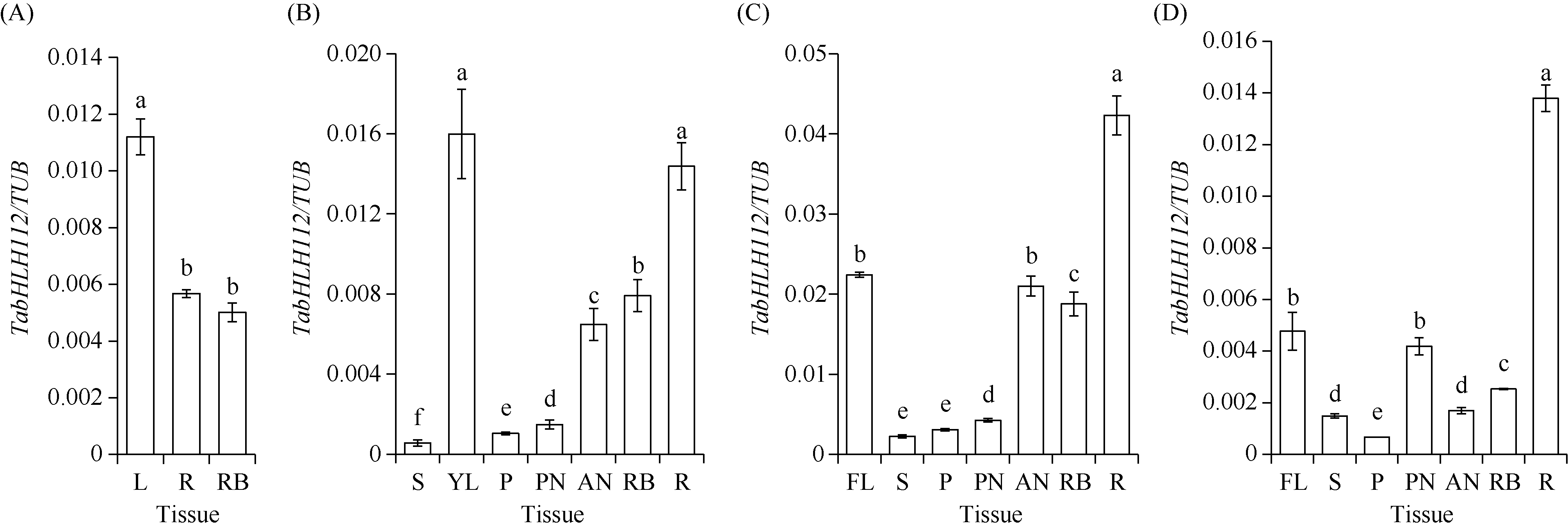
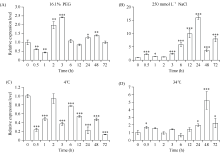
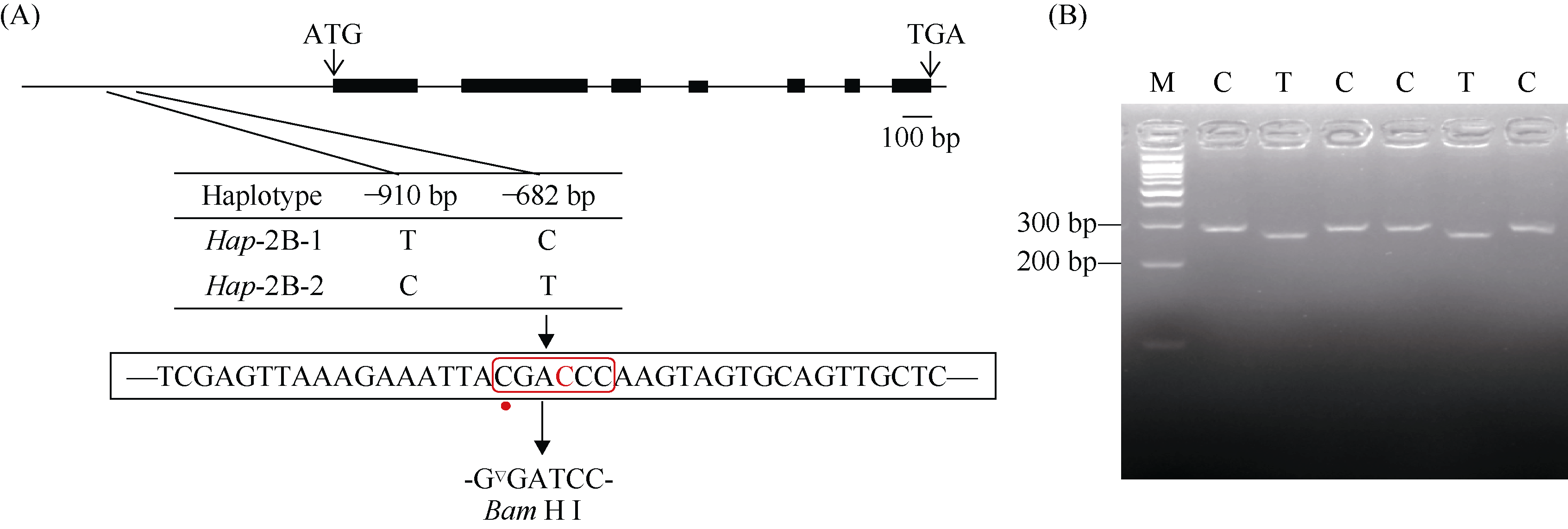
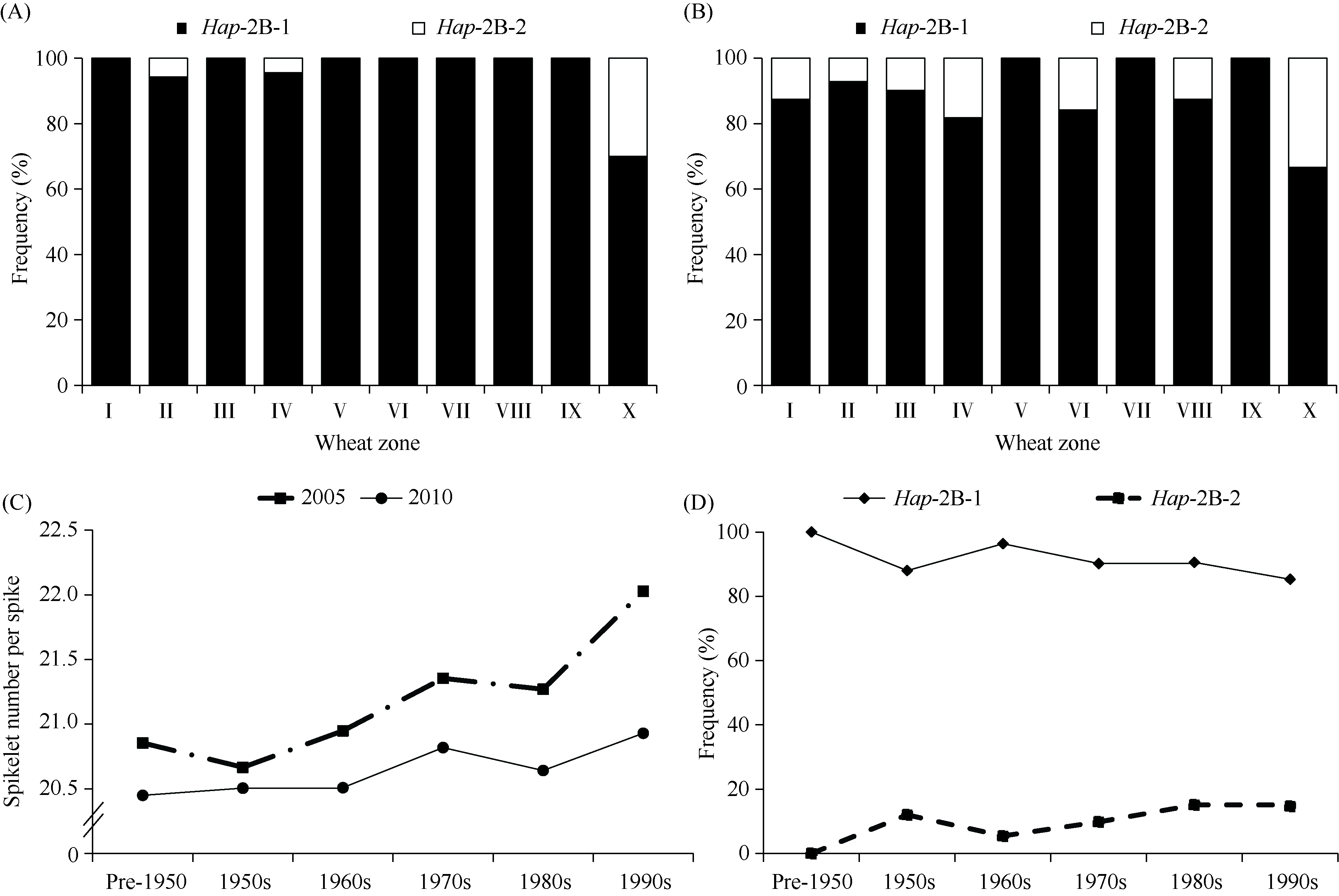
bHLH (basic Helix-Loop-Helix)转录因子在植物生长发育过程中发挥着重要作用。本研究克隆了小麦TabHLH112-2B, 该基因由7个外显子和6个内含子构成, 编码444个氨基酸, 在315~364位氨基酸处具有一个典型的HLH保守结构域。组织表达模式分析表明TabHLH112-2B在小麦幼苗期、拔节期、抽穗期和开花期的各个组织中均有表达, 其中在叶和根中表达量较高。顺式作用元件分析发现TabHLH112-2B启动子区含有植物激素应答、胁迫响应、与分生组织发育相关的多种顺式作用元件, qRT-PCR结果显示其表达响应ABA、IAA、MeJA等植物激素和干旱、高盐、低温、高温等胁迫处理。基因组序列多态性分析检测到启动子区域的2个SNP, 分为2种单倍型。根据SNP-682开发分子标记并进行关联分析, 发现该标记与干旱、高温等多种环境下的每穗小穗数显著相关, Hap-2B-2为每穗小穗数多的优异单倍型。研究结果为培育高产广适小麦新品种的分子标记辅助育种提供了优异基因资源和技术支撑。
| [1] | Dowla U M, Edwards I, O’Hara G, Islam S, Ma W. Developing wheat for improved yield and adaptation under a climate: optimization of a few key genes. Engineering, 2018, 4: 183-202. |
| [2] | 卢红芳. 高温、干旱及其复合胁迫对小麦籽粒谷蛋白大聚合体、淀粉粒度分布和品质性状的影响. 河南农业大学博士学位论文, 河南郑州, 2013. |
| Lu H F. Effects of High Temperature, Drought and Composite Stress on Large Aggregate, Starch Size Distribution and Quality Traits of Wheat Grain. PhD Dissertation of Henan Agricultural University, Zhengzhou, Henan, China, 2013 (in Chinese with English abstract). | |
| [3] |
Li Y P, Li L, Zhao M, Guo L, Guo X, Zhao D, Batool A, Dong B D, Xu H X, Cui S J, Zhang A M, Fu X D, Li J M, Jing R L, Liu X G. Wheat FRIZZY PANICLE activates VERNALIZATION1-A and HOMEOBOX4-A to regulate spike development in wheat. Plant Biotechnol J, 2021, 19: 1141-1154.
doi: 10.1111/pbi.v19.6 |
| [4] |
Ma J, Ding P Y, Liu J J, Li T, Zou Y Y, Habib A, Mu Y, Tang H P, Jiang Q T, Liu Y X, Chen G Y, Wang J R, Deng M, Qi P F, Li W, Pu Z E, Zheng Y L, Wei Y M, Lan X J. Identification and validation of a major and stably expressed QTL for spikelet number per spike in bread wheat. Theor Appl Genet, 2019, 132: 3155-3167.
doi: 10.1007/s00122-019-03415-z pmid: 31435704 |
| [5] |
Ding P Y, Mo Z Q, Tang H P, Mu Y, Deng M, Jiang Q T, Liu Y X, Chen G D, Chen G Y, Wang J R. A major and stable QTL for wheat spikelet number per spike validated in different genetic backgrounds. J Integr Agric, 2022, 21: 1551-1562.
doi: 10.1016/S2095-3119(20)63602-4 |
| [6] | Kuzay S, Lin H Q, Li C X, Chen S S, Woods D P, Zhang J L, Lan T Y, Korff M V, Dubcovsky J. WAPO-A1 is the causal gene of the 7AL QTL for spikelet number per spike in wheat. PLoS Genet, 2022, 18: 1009747. |
| [7] |
Wittern L M, Barrero J M, Bovill W D, Verbyla K L, Hughes T, Swain S M, Steed G, Webb A A R, Gardner K, Greenland A, Jacobs J, Frohberg C, Schmidt R C, Cavanagh C, Rohde A, Davey M W, Hannah M A. Overexpression of the WAPO-A1 gene increases the number of spikelets per spike in bread wheat. Sci Rep, 2022, 12: 14229.
doi: 10.1038/s41598-022-18614-w pmid: 35987959 |
| [8] |
Jin J P, Zhang H, Kong L, Gao G, Luo J C. PlantTFDB 3.0: a portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res, 2014, 42: D1182-1187.
doi: 10.1093/nar/gkt1016 |
| [9] |
Liu Y J, Ji X Y, Nie X G, Qu M, Zheng L, Tan Z L, Zhao H M, Huo L, Liu S N, Zhang B, Wang Y C. Arabidopsis AtbHLH112 regulates the expression of genes involved in abiotic stress tolerance by binding to their E-box and GCG-box motifs. New Phytol, 2015, 207: 692-709.
doi: 10.1111/nph.2015.207.issue-3 |
| [10] |
Castilhos G, Lazzarotto F, Spagnolo-Fonini L, Bodanese- Zanettini M H, Margis-Pinheiro M. Possible roles of basic helix-loop-helix transcription factors in adaptation to drought. Plant Sci, 2014, 223: 1-7.
doi: 10.1016/j.plantsci.2014.02.010 pmid: 24767109 |
| [11] |
Gu X Y, Gao S X, Li J, Song P Y, Zhang Q, Guo J F, Wang X Y, Han X Y, Wang X J, Zhu Y, Zhu Z G. The bHLH transcription factor regulated gene OsWIH2 is a positive regulator of drought tolerance in rice. Plant Physiol Biochem, 2021, 169: 269-279.
doi: 10.1016/j.plaphy.2021.11.031 |
| [12] | Zhang L, Xiang Z P, Li J F, Wang S Y, Chen Y, Liu Y, Mao D D, Luan S, Chen L B. bHLH57 confers chilling tolerance and grain yield improvement in rice. Plant Cell Environ, 2022, doi: 10.1111/pce.14513. |
| [13] |
Liu H, Yang Y, Liu D D, Wang X Y, Zhang L S. Transcription factor TabHLH49 positively regulates dehydrin WZY2 gene expression and enhances drought stress tolerance in wheat. BMC Plant Biol, 2020, 20: 259.
doi: 10.1186/s12870-020-02474-5 |
| [14] |
Guo X J, Fu Y X, Lee Y J, Chern M S, Li M L, Cheng M P, Dong H X, Yuan Z W, Gui L X, Yin J X, Qing H, Zhang C B, Pu Z E, Liu Y J, Li W T, Li W, Qi P F, Chen G Y, Jiang Q T, Ma J, Chen X W, Wei Y M, Zheng Y L, Wu Y R, Liu B, Wang J R. The PGS1 basic helix-loop-helix protein regulates Fl3 to impact seed growth and grain yield in cereals. Plant Biotechnol J, 2022, 20: 1311-1326.
doi: 10.1111/pbi.v20.7 |
| [15] |
Li L, Mao X G, Wang J Y, Chang X P, Reynolds M, Jing R L. Genetic dissection of drought and heat-responsive agronomic traits in wheat. Plant Cell Environ, 2019, 42: 2540-2553.
doi: 10.1111/pce.v42.9 |
| [16] |
Li L, Peng Z, Mao X G, Wang J Y, Chang X P, Reynolds M, Jing R L. Genome-wide association study reveals genomic regions controlling root and shoot traits at late growth stages in wheat. Ann Bot, 2019, 124: 993-1006.
doi: 10.1093/aob/mcz041 |
| [17] |
Hao C Y, Wang L F, Ge H M, Dong Y C, Zhang X Y. Genetic diversity and linkage disequilibrium in Chinese bread wheat (Triticum aestivum L.) revealed by SSR markers. PLoS One, 2011, 6: e17279.
doi: 10.1371/journal.pone.0017279 |
| [18] |
Hao Y Q, Zong X M, Ren P, Qian Y Q, Fu A G. Basic Helix-Loop-Helix (bHLH) transcription factors regulate a wide range of functions in Arabidopsis. Int J Mol Sci, 2021, 22: 7152.
doi: 10.3390/ijms22137152 |
| [19] |
Yang T R, Yao S F, Hao L, Zhao Y Y, Lu W J, Xiao K. Wheat bHLH-type transcription factor gene TabHLH1 is crucial in mediating osmotic stresses tolerance through modulating largely the ABA-associated pathway. Plant Cell Rep, 2016, 35: 2309-2323.
doi: 10.1007/s00299-016-2036-5 |
| [20] |
Wang R, Yu M M, Xia J Q, Xing J P, Fan X P, Xu Q H, Cang J, Zhang D. Overexpression of TaMYC2 confers freeze tolerance by ICE-CBF-COR module in Arabidopsis thaliana. Front Plant Sci, 2022, 13: 1042889.
doi: 10.3389/fpls.2022.1042889 |
| [21] |
Zhao F N, Lei J, Wang R Y, Zhang Q, Qi Y, Zhang K, Guo Q, Wang H L. Environmental determination of spring wheat yield in a climatic transition zone under global warming. Int J Biometeorol, 2022, 66: 481-491.
doi: 10.1007/s00484-021-02196-9 |
| [22] | 袁蕊, 李萍, 胡晓雪, 宗毓铮, 孙敏, 董琦, 郝兴宇. 干旱胁迫对小麦生理特性及产量的影响. 山西农业科学, 2016, 44: 1446-1449. |
| Yuan R, Li P, Hu X X, Zong Y Z, Sun M, Dong Q, Hao X Y. Effects of drought stress on physiological characteristics and yield of wheat. J Shanxi Agri Sci, 2016, 44: 1446-1449 (in Chinese with English abstract). | |
| [23] |
杨绚, 汤绪, 陈葆德, 田展, 钟洪麟. 气候变暖背景下高温胁迫对中国小麦产量的影响. 地理科学进展, 2013, 32: 1771-1779.
doi: 10.11820/dlkxjz.2013.12.006 |
| Yang X, Tang X, Chen B D, Tian Z, Zhong H L. Effects of high temperature stress on wheat yield in China under the background of climate warming. Prog Geog, 2013, 32: 1771-1779 (in Chinese with English abstract). | |
| [24] |
Itam M O, Mega R, Gorafi Y S A, Yamasaki Y, Tahir I S A, Akashi K, Tsujimoto H. Genomic analysis for heat and combined heat-drought resilience in bread wheat under field conditions. Theor Appl Genet, 2022, 135: 337-350.
doi: 10.1007/s00122-021-03969-x |
| [25] |
Hammond-Kosack M C U, King R, Kanyuka K, Hammond-Kosack K E. Exploring the diversity of promoter and 5'UTR sequences in ancestral, historic and modern wheat. Plant Biotechnol J, 2021, 19: 2469-2487.
doi: 10.1111/pbi.13672 pmid: 34289221 |
| [26] |
Xia C, Zhang L C, Zou C, Gu Y Q, Duan J L, Zhao G Y, Wu J J, Liu Y, Fang X H, Gao L F, Jiao Y N, Sun J Q, Pan Y H, Liu X, Jia J Z, Kong X Y. A TRIM insertion in the promoter of Ms2 causes male sterility in wheat. Nat Commun, 2017, 8: 15407.
doi: 10.1038/ncomms15407 |
| [27] |
Li C N, Wang J Y, Li L, Li J L, Zhuang M J, Li B, Li Q R, Huang J F, Du Y, Wang J P, Fan Z P, Mao X G, Jing R L. TaMOR is essential for root initiation and improvement of root system architecture in wheat. Plant Biotechnol J, 2022, 20: 862-875.
doi: 10.1111/pbi.v20.5 |
| [28] |
Xue Y H, Wang J Y, Mao X G, Li C N, Li L, Yang X, Hao C Y, Chang X P, Li R Z, Jing R L. Association analysis revealed that TaPYL4genes are linked to plant growth related traits in multiple environment. Front Plant Sci, 2021, 12: 641087.
doi: 10.3389/fpls.2021.641087 |
| [29] |
Hu J, Jin Q, Ma Y P. AfLFY, a LEAFY homolog in Argyranthemum frutescens, controls flowering time and leaf development. Sci Rep, 2020, 10: 1616.
doi: 10.1038/s41598-020-58570-x |
| [30] |
Sablowski R. Flowering and determinacy in Arabidopsis. J Exp Bot, 2007, 58: 899-907.
doi: 10.1093/jxb/erm002 pmid: 17293602 |
| [31] |
Weigel D, Nilsson O. A developmental switch sufficient for flower initiation in diverse plants. Nature, 1995, 377: 495-500.
doi: 10.1038/377495a0 |
| [1] | 郝倩琳, 杨廷志, 吕新茹, 秦慧敏, 王亚林, 贾晨飞, 夏先春, 马武军, 徐登安. 小麦胚芽鞘长度QTL定位和GWAS分析[J]. 作物学报, 2024, 50(3): 590-602. |
| [2] | 琚吉浩, 马超, 王添宁, 吴毅, 董钟, 方美娥, 陈钰姝, 张均, 付国占. 小麦TaPOD家族的全基因组鉴定及表达分析[J]. 作物学报, 2024, 50(3): 779-792. |
| [3] | 王琼, 朱宇翔, 周密密, 张威, 张红梅, 陈新, 陈华涛, 崔晓艳. 大豆叶型性状全基因组关联分析与候选基因鉴定[J]. 作物学报, 2024, 50(3): 623-632. |
| [4] | 赵荣荣, 丛楠, 赵闯. 基于Landsat 8影像提取豫中地区冬小麦和夏玉米分布信息的最佳时相选择[J]. 作物学报, 2024, 50(3): 721-733. |
| [5] | 张宝华, 刘佳静, 田晓, 田旭钊, 董阔, 武郁洁, 肖凯, 李小娟. 小麦TaSPX1基因的克隆、表达及耐低氮逆境的功能研究[J]. 作物学报, 2024, 50(3): 576-589. |
| [6] | 柯会锋, 苏红梅, 孙正文, 谷淇深, 杨君, 王国宁, 徐东永, 王洪这, 吴立强, 张艳, 张桂寅, 马峙英, 王省芬. 棉花现代品种资源产量与纤维品质性状鉴定及分子标记评价[J]. 作物学报, 2024, 50(2): 280-293. |
| [7] | 田春艳, 边芯, 郎荣斌, 俞华先, 桃联安, 安汝东, 董立华, 张钰, 经艳芬. 甘蔗3个育种性状与SSR标记的关联分析及优异等位变异发掘[J]. 作物学报, 2024, 50(2): 310-324. |
| [8] | 陈天, 李昱樱, 荣二花, 吴玉香. 棉属人工异源四倍体后代性状鉴定及花器转录组学分析[J]. 作物学报, 2024, 50(2): 325-339. |
| [9] | 马娟, 曹言勇. 玉米杂交群体产量性状及其特殊配合力全基因组关联分析[J]. 作物学报, 2024, 50(2): 363-372. |
| [10] | 张康, 聂志刚, 王钧, 李广. 温度升高下APSIM模型春小麦籽粒生长参数敏感性分析及优化[J]. 作物学报, 2024, 50(2): 464-477. |
| [11] | 谭丹, 陈家婷, 郜钰, 张晓军, 李欣, 闫贵云, 李锐, 陈芳, 常利芳, 张树伟, 郭慧娟, 畅志坚, 乔麟轶. 小麦穗型相关生长素通路基因发掘及TaARF23-A与小穗数关联分析[J]. 作物学报, 2024, 50(2): 506-513. |
| [12] | 李艳, 方宇辉, 王永霞, 彭超军, 华夏, 齐学礼, 胡琳, 许为钢. 不同磷胁迫处理转OsPHR2小麦的转录组学分析[J]. 作物学报, 2024, 50(2): 340-353. |
| [13] | 谢炜, 贺鹏, 马宏亮, 雷芳, 黄秀兰, 樊高琼, 杨洪坤. 秋闲期秸秆覆盖与施磷对冬小麦氮素吸收利用的影响[J]. 作物学报, 2024, 50(2): 440-450. |
| [14] | 李俣佳, 许豪, 于士男, 唐建卫, 李巧云, 高艳, 郑继周, 董纯豪, 袁雨豪, 郑天存, 殷贵鸿. 小麦骨干亲本周8425B抗条锈病优异基因在其衍生品种中的遗传解析[J]. 作物学报, 2024, 50(1): 16-31. |
| [15] | 黄钰杰, 张啸天, 陈会丽, 王宏伟, 丁双成. 玉米ZmC2s基因家族鉴定及ZmC2-15耐热功能分析[J]. 作物学报, 2023, 49(9): 2331-2343. |
|