欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (10): 1604-1612.doi: 10.3724/SP.J.1006.2019.81091
• 研究简报 • 上一篇
周萍萍1,2,颜红海1,2,3,*( ),彭远英2,*()
),彭远英2,*()
ZHOU Ping-Ping1,2,YAN Hong-Hai1,2,3,*(),PENG Yuan-Ying2,*()
摘要:
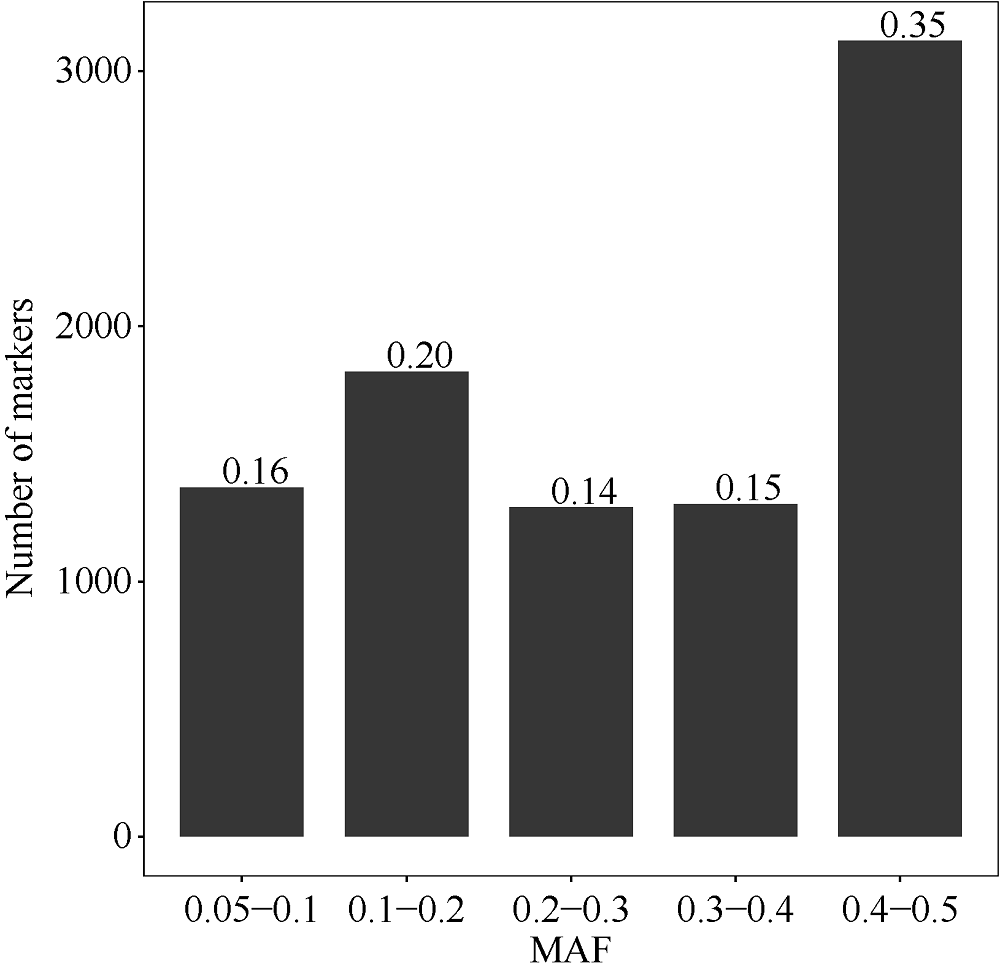
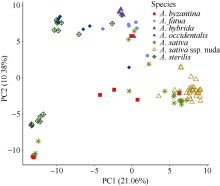
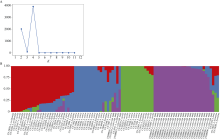
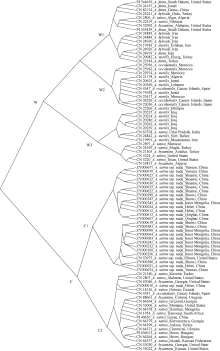
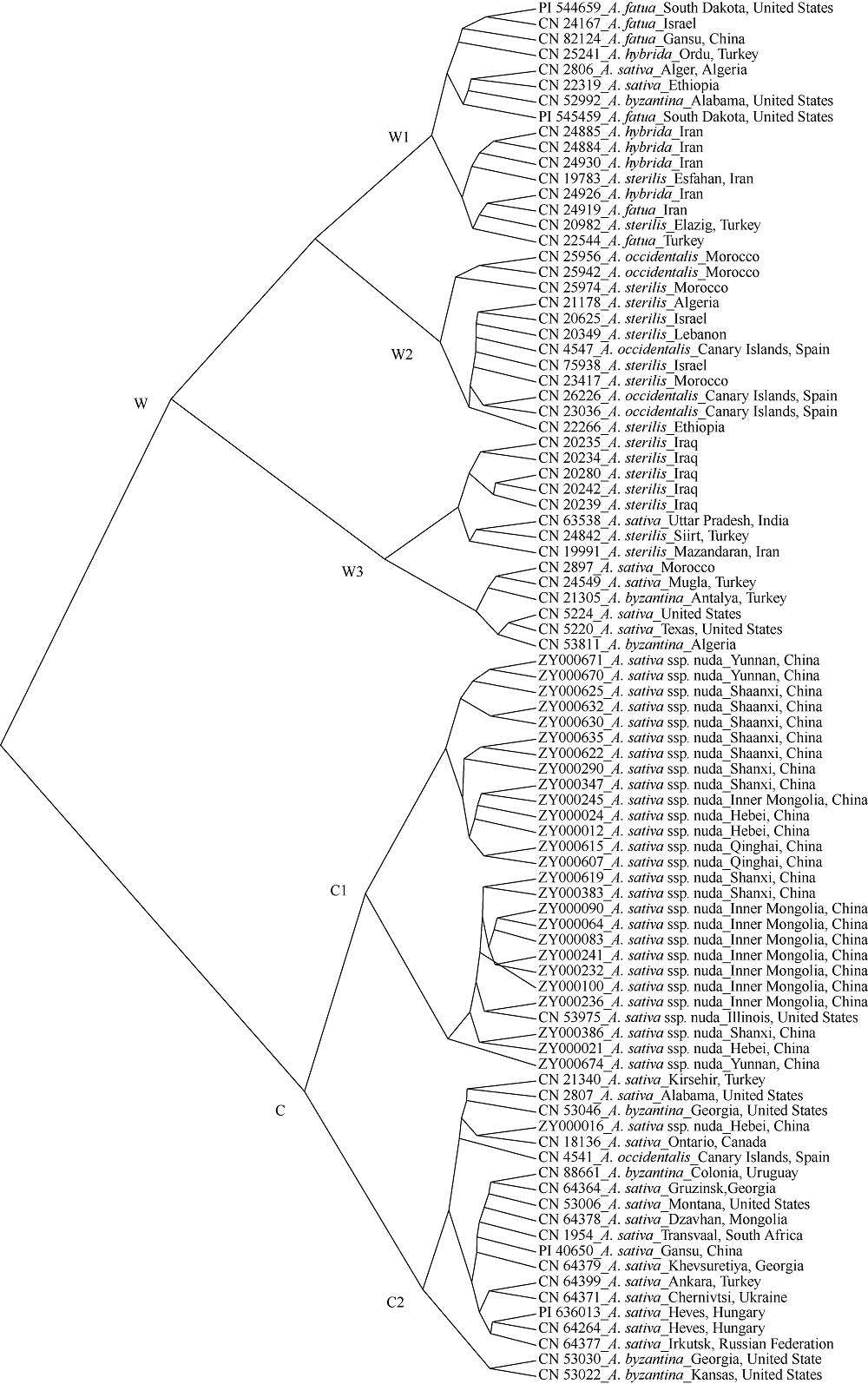
栽培六倍体燕麦是世界重要粮食作物, 理清其起源对燕麦种质资源的高效利用和保护具有重要意义。本研究利用GBS (genotyping by sequencing)对27份来自中国的大粒裸燕麦材料测序, 结合先前发表的包括6个六倍体燕麦种在内的66份燕麦材料的GBS数据进行SNP挖掘。UNEAK管道挖掘共计得到MAF大于0.5, call rate大于0.95的SNP标记8902个。进一步剔除缺失值大于0.15的4个燕麦材料后, 对其余89份材料进行PCA分析、STRUCTURE分析以及UPGMA聚类分析。结果表明, 在野生种中, 除A. sterilis外, 大多数来自同一物种的材料聚为一类, 不同物种间能够较好地分开, 表明这些物种之间存在较强的遗传分化。聚类分析将供试材料分为分别代表野生种和栽培种的2支, 表明野生种和栽培种之间存在明显的遗传差异; 在栽培种中, A. sativa与A. byzantina具有较高的遗传多样性, 分散在不同的类群中, 二者未出现明显的遗传分化, 具有较高的遗传同质性, A. sativa ssp. nuda与A. sativa亲缘关系较近, 但存在一定的遗传分化, 因此形成独立的类群。值得注意的是, 来自野生种A. sterilis的材料被分在2个类群中, 其中来自西南亚地区(伊朗-伊拉克-土耳其地区)的居群与A. sativa和A. byzantina聚在一起, 揭示此地区的A. sterilis居群可能是A. sativa和A. byzantina的祖先种。野生种A. hybrida显示出与A. fatua较高的遗传同质性, 因此将其作为A. fatua的亚种较为合理。本研究为栽培六倍体燕麦起源提供了理论依据。
| [1] | Kong L, Huo H, Mao P . Antioxidant response and related gene expression in aged oat seed. Front Plant Sci, 2015,6:158 |
| [2] | FAOSTATS. . 2016 |
| [3] | Coffman F A . Oat History, Identification and Classification. Washington D. C: US Department of Agriculture, Agricultural Research Service. 1977. pp 3-30 |
| [4] | Cox D J, Frey K J . Improving cultivated oats ( Avena sativa L.) with alleles for vegetative growth index from A. sterilis L. Theor Appl Genet, 1984,68:239-245 |
| [5] | Branson C V, Frey K J . Recurrent selection for groat oil content in oat. Crop Sci, 1989,29:1382-1387 |
| [6] | Zamir D . Improving plant breeding with exotic genetic libraries. Nat Rev Genet, 2001,2:983-989 |
| [7] | Loskutov I G, Rines H W. Avena. In: Kole C, ed. Wild Crop Relatives: Genomic and Breeding Resources. Heidelberg: Springer Press, 2011. pp 109-183 |
| [8] | Baum B R. Oats: Wild and Cultivated. A Monograph of the Genus Avena L. (Poaceae). Ottawa: Minister of Supply and Services Press, 1977. |
| [9] | Ladizinsky G. Studies in Oat Evolution. A Man’s Life with Avena (Springer Briefs in Agriculture). Heidelberg: Springer Press, 2012. pp 1-18 |
| [10] | Coffman F A . Origin of cultivated oats. J Am Soc Agron, 1946,38:983-1002 |
| [11] | 郑殿升, 张宗文 . 大粒裸燕麦(莜麦)(Avena nuda L.)起源及分类问题的探讨. 植物遗传资源学报, 2011,5:667-670. |
| Zheng D S, Zhang Z W . Discussion on the origin and taxonomy of naked oat ( Avena nuda L.). J Plant Genet Resour, 2011,5:667-670 (in Chinese with English abstract) | |
| [12] | Zhou X, Jellen E N, Murphy J P . Progenitor germplasm of domisticated hexaploid oat. Crop Sci, 1999,39:1208-1214 |
| [13] | Loskutov I G . On evolutionary pathways of Avena species. Genet Resour Crop Evol, 2008,55:211-220 |
| [14] | 刘青, 刘欢, 林磊 . 燕麦属系统学研究进展. 热带亚热带植物学报, 2014,5:516-524 |
| Liu Q, Liu H, Lin L . Research advances on systematics of Avena( Pooideae, Poaceae). J Trop Subtrop Bot, 2014,5:516-524 (in Chinese with English abstract) | |
| [15] | Ladizinsky G. The domestication and history of oats. In: Mattsson B, Lyhagen R, eds. Proceedings of the 3rd International Oat Conference, Lund, Sweden. Svalof AB, 1988. pp 7-12 |
| [16] | Baum B R . Extrapolation of the predomesticated hexaploid cultivated oats. Evolution, 1973,27:518-523 |
| [17] | Huang Y F, Poland J A, Wight C P, Tinker N A . Using genotyping-by-sequencing (GBS) for genomic discovery in cultivated oat. PLoS One, 2014,9:e102448 |
| [18] | Chew P, Meade K, Hayes A, Harjes C, Bao Y, Beattie A D, Puddephat I, Gusmini G, Tanksley S D . A study on the genetic relationships of Avena taxa and the origins of hexaploid oat. Theor Appl Genet, 2016,129:1405-1415 |
| [19] | Yan H, Bekele W A, Wight C P, Peng Y, Langdon T, Latta R G, Fu Y B, Diederichsen A, Howarth C J, Jellen E N, Boyle B, Wei Y, Tinker N A . High-density marker profiling confirms ancestral genomes of Avena species and identifies D-genome chromosomes of hexaploid oat. Theor Appl Genet, 2016,129:2133-2149 |
| [20] | Chaffin A S, Huang Y F, Smith S, Bekele W A, Babiker E, Gnanesh B N, Foresman B J, Blanchard S G, Jay J J, Reid R W, Wight C P, Chao S, Islamovic E, Kolb F L, McCartney C, Mitchell F J W, Beattie A D, Bjornstad A, Bonman J M, Langdon T, Howarth C J, Brouwer C R, Jellen E N, Klos K E, Poland J A, Hsieh T F, Brown R, Jackson E, Schlueter J A, Tinker N A . A consensus map in cultivated hexaploid oat reveals conserved grass synteny with substantial subgenome rearrangement. Plant Genome, 2016,9. doi: 10.3835/plantgenome2015.10.0102. |
| [21] | Torkamaneh D, Laroche J, Belzile F . Genome-wide SNP calling from genotyping by sequencing (GBS) data: a comparison of seven pipelines and two sequencing technologies. PLoS One, 2016,11:e0161333 |
| [22] | Lu F, Lipka A E, Glaubitz J, Elshire R, Cherney J H, Casler M D, Buckler E S, Costich D E . Switchgrass genomic diversity, ploidy, and evolution: novel insights from a Nnetwork-based SNP discovery protocol. PLoS Genet, 2013,9:139-147 |
| [23] | Bradbury P J, Zhang Z, Kroon D E, Casstevens T M, Ramdoss Y, Buckler E S . TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007,23:2633-2635 |
| [24] | Pritchard J, Stephens M, Donnelly . Inference of population structure using multilocus genotype data. Genetics, 2000,155:945-959 |
| [25] | Evanno G, Regnaut S, Goudet J . Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol, 2005,14:2611-2620 |
| [26] | Earl D A, Vonholdt B M . STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour, 2012,4:359-361 |
| [27] | R Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. . 2016. |
| [28] | Souza E, Sorrells M E . Relationships among 70 north American oat germplasms: I. cluster analysis using quantitative characters. Crop Sci, 1991,31:599-605 |
| [29] | Koch. K . Beiträge zu einer flora des orients. Linnaeus, 1948,21:289-443 |
| [30] | Baohong G, Zhou X, Murphy J P . Genetic variation within Chinese and Western cultivated oat accessions. Cereal Res Commun, 2003,31:339-346 |
| [31] | Malzew A . Wild and cultivated oats: Sectio Eu Avena Griseb. Bull Appl Bot, Genet Plant Breed, 1930,38:473-506 (in Russian with English abstract) |
| [1] | 刘丹, 周彩娥, 王晓婷, 吴启蒙, 张旭, 王琪琳, 曾庆东, 康振生, 韩德俊, 吴建辉. 利用集群分离分析结合高密度芯片快速定位小麦成株期抗条锈病基因YrC271[J]. 作物学报, 2022, 48(3): 553-564. |
| [2] | 郑向华, 叶俊华, 程朝平, 魏兴华, 叶新福, 杨窑龙. 利用SNP标记进行水稻品种籼粳鉴定[J]. 作物学报, 2022, 48(2): 342-352. |
| [3] | 张春, 赵小珍, 庞承珂, 彭门路, 王晓东, 陈锋, 张维, 陈松, 彭琦, 易斌, 孙程明, 张洁夫, 傅廷栋. 甘蓝型油菜千粒重全基因组关联分析[J]. 作物学报, 2021, 47(4): 650-659. |
| [4] | 孙程明,陈锋,陈松,彭琦,张维,易斌,张洁夫,傅廷栋. 甘蓝型油菜每角粒数的全基因组关联分析[J]. 作物学报, 2020, 46(01): 147-153. |
| [5] | 孙程明,陈松,彭琦,张维,易斌,张洁夫,傅廷栋. 甘蓝型油菜角果长度性状的全基因组关联分析[J]. 作物学报, 2019, 45(9): 1303-1310. |
| [6] | 杨芳萍,刘金栋,郭莹,贾奥琳,闻伟鄂,巢凯翔,伍玲,岳维云,董亚超,夏先春. 普通小麦‘Holdfast’条锈病成株抗性QTL定位[J]. 作物学报, 2019, 45(12): 1832-1840. |
| [7] | 白彦明,李龙,王绘艳,柳玉平,王景一,毛新国,昌小平,孙黛珍,景蕊莲. 蚂蚱麦和小白麦衍生系的遗传多样性分析[J]. 作物学报, 2019, 45(10): 1468-1477. |
| [8] | 张学勇,马琳,郑军. 作物驯化和品种改良所选择的关键基因及其特点[J]. 作物学报, 2017, 43(02): 157-170. |
| [9] | 陈广凤,陈建省,田纪春. 小麦株高相关性状与SNP标记全基因组关联分析[J]. 作物学报, 2015, 41(10): 1500-1509. |
| [10] | 肖永贵, 李思敏, 李法计, 张宏燕, 陈新民, 王德森, 夏先春, 何中虎. 两种施肥环境下冬小麦京411及其衍生系产量和生理性状的遗传分析[J]. 作物学报, 2015, 41(09): 1333-1342. |
| [11] | 邱先进,袁志华,陈凯,杜斌,何文静,杨隆维,徐建龙,邢丹英,吕文恺. 用全基因组关联分析解析籼稻垩白的遗传基础[J]. 作物学报, 2015, 41(07): 1007-1016. |
| [12] | 周艳华,曹红利,岳川,王璐,郝心愿,王新超*,杨亚军*. 冷驯化不同阶段茶树DNA甲基化模式的变化[J]. 作物学报, 2015, 41(07): 1047-1055. |
| [13] | 肖炳光, 邱杰,曹培健,桂毅杰,卢秀萍,李永平,樊龙江. 利用基因组简约法开发烟草SNP标记及遗传作图[J]. 作物学报, 2014, 40(03): 397-404. |
| [14] | 李飞,徐建飞,刘杰,段绍光,卞春松,Jiwan P. PALTA,金黎平. 三个耐冻性不同的马铃薯野生种中FAD2基因的克隆及表达分析[J]. 作物学报, 2014, 40(01): 45-53. |
| [15] | 陶爱芬, 祁建民, 粟建光, 李爱青, 李木兰, 方平平, 林荔辉, 徐建堂, 吴建梅, 林培清. SRAP和ISSR及两种方法结合在分析黄麻属起源与演化上的比较[J]. 作物学报, 2011, 37(12): 2277-2284. |
|
||