欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (9): 1368-1379.doi: 10.3724/SP.J.1006.2020.94188
陈淼( ), 谢赛, 王超智, 李焱龙, 张献龙, 闵玲*()
), 谢赛, 王超智, 李焱龙, 张献龙, 闵玲*()
CHEN Miao(), XIE Sai, WANG Chao-Zhi, LI Yan-Long, ZHANG Xian-Long, MIN Ling*()
摘要:
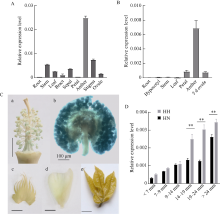
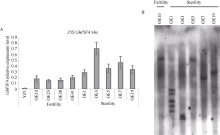
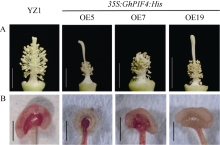
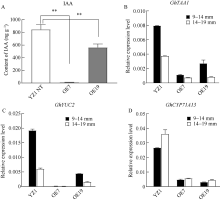
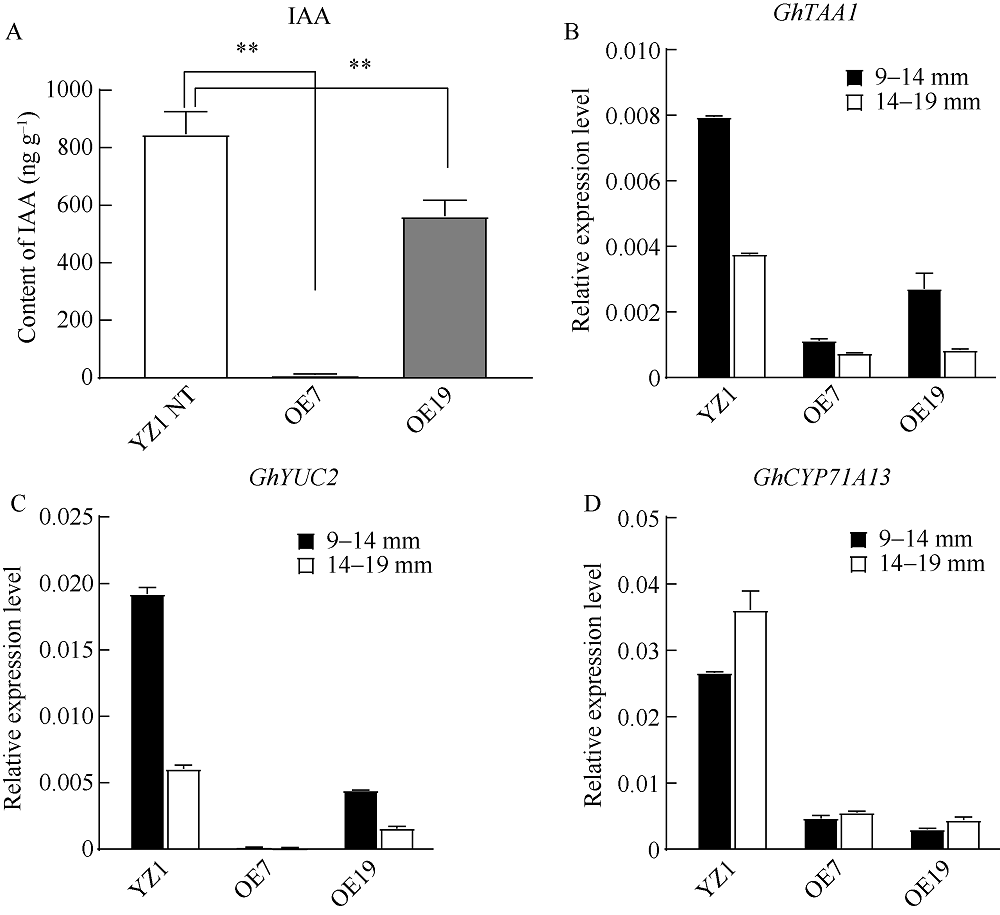
PIF4属于PIF家族(PHYTOCHROME INTERACTING FACTORS FAMILY), 是一个响应温度变化的关键性转录因子, 广泛参与植物热信号传导和激素信号通路。本研究从陆地棉‘YZ1’中克隆获得GhPIF4基因, 亚细胞定位结果表明, GhPIF4是一个核蛋白。qRT-PCR分析和proGhPIF4:GUS转基因棉花GUS染色结果显示, GhPIF4基因在棉花花药中高量表达, 且其表达在高温敏感型材料‘H05’中受到高温的显著诱导。在棉花中超量表达GhPIF4, 不同的转基因株系表现不同的育性, 表达量高的3个转基因系(OE5、OE7和OE19)开花当天的花药不开裂, 花粉的活力均低于对照‘YZ1’, 而超表达程度略低的OE10, 花药正常开裂, 花粉活性正常, 表明GhPIF4导致的花药败育具有剂量效应。对OE7和OE19在9~14 mm (绒毡层降解期)、14~19 mm (有丝分裂I期)中生长素的含量以及生长素合成关键基因GhTAA1、GhYUC2、GhCYP71A13的表达分析发现基因的表达与生长素含量下降变化一致。推测超表达GhPIF4在花药后期可能部分模拟棉花的高温响应状态, 而组成型超表达GhPIF4可能改变了营养器官或早期花药中生长素的含量, 亦或花药中生长素含量过低也会导致花药败育。以上结果为深入解析GhPIF4基因功能及了解高温胁迫下棉花花药败育的机制提供参考。
| [1] |
Zhang Q, Shen B Z, Dai X K, Mei M H, Saghai Maroof M A, Li Z B. Using bulked extremes and recessive class to map genes for photoperiod-sensitive genic male sterility in rice. Proc Natl Acad Sci USA, 1994,91:8675-8679.
pmid: 7915844 |
| [2] |
Chen L, Liu Y G. Male sterility and fertility restoration in crops. Annu Rev Plant Biol, 2014,65:579-606.
doi: 10.1146/annurev-arplant-050213-040119 pmid: 24313845 |
| [3] | 袁隆平. 选育水稻光、温敏核不育系的技术策略. 杂交水稻, 1992, (1):1-4. |
| Yuan L P. Technical strategy for breeding rice light and temperature sensitive genic male sterile lines. Hybrid Rice, 1992, (1):1-4 (in Chinese). | |
| [4] |
Endo M, Tsuchiya T, Hamada K, Kawamura S, Yano K, Ohshima M, Higashitani A, Watanabe M, Kawagishi-Kobayashi M. High temperatures cause male sterility in rice plants with transcriptional alterations during pollen development. Plant Cell Physiol, 2009,50:1911-1922.
doi: 10.1093/pcp/pcp135 pmid: 19808807 |
| [5] |
Zinn K E, Tunc-Ozdemir M, Harper J F. Temperature stress and plant sexual reproduction: uncovering the weakest links. J Exp Bot, 2010,61:1959-1968.
pmid: 20351019 |
| [6] |
Dong B, Zheng X, Liu H P, Able J A, Yang H, Zhao H, Zhang M M, Qiao Y Z, Wang Y K, Liu M Y. Effects of drought stress on pollen sterility, grain yield, abscisic acid and protective enzymes in two winter wheat cultivars. Front Plant Sci, 2017,8:1008.
pmid: 28676806 |
| [7] |
De Storme N, Geelen D. The impact of environmental stress on male reproductive development in plants: biological processes and molecular mechanisms. Plant Cell Environ, 2014,37:1-18.
doi: 10.1111/pce.12142 pmid: 23731015 |
| [8] |
Khanna R, Huq E, Kikis E A, Al-Sady B, Lanzatella C, Quail P H. A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix- loop-helix transcription factors. Plant Cell, 2004,16:3033-3044.
doi: 10.1105/tpc.104.025643 pmid: 15486100 |
| [9] |
Ni M, Tepperman J M, Quail P H. Binding of phytochrome B to its nuclear signalling partner PIF3 is reversibly induced by light. Nature, 1999,400:781-784.
doi: 10.1038/23500 pmid: 10466729 |
| [10] |
Huq E, Al-Sady B, Hudson M, Kim C, Apel K, Quail P H. PHYTOCHROME-INTERACTING FACTOR 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science, 2004,305:1937-1941.
pmid: 15448264 |
| [11] |
Huq E, Quail P H. PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis. EMBO J, 2002,21:2441-2450.
doi: 10.1093/emboj/21.10.2441 pmid: 12006496 |
| [12] |
Leivar P, Monte E, Al-Sady B, Carle C, Storer A, Alonso J M, Ecker J R, Quail P H. The Arabidopsis phytochrome-interacting factor PIF7, together with PIF3 and PIF4, regulates responses to prolonged red light by modulating phyB levels. Plant Cell, 2008,20:337-352.
pmid: 18252845 |
| [13] |
Quint M, Delker C, Franklin K A, Wigge P A, Halliday K J, van Zanten M. Molecular and genetic control of plant thermomorphogenesis. Nat Plants, 2016,2:15190.
doi: 10.1038/nplants.2015.190 pmid: 27250752 |
| [14] |
Casal J J, Balasubramanian S. Thermomorphogenesis. Annu Rev Plant Biol, 2019,70:321-346.
pmid: 30786235 |
| [15] |
Legris M, Klose C, Burgie E S, Rojas C C R, Neme M, Hiltbrunner A, Wigge P A, Schäfer E, Vierstra R D, Casal J J. Phytochrome B integrates light and temperature signals in Arabidopsis. Science, 2016,354:897-900.
pmid: 27789798 |
| [16] |
Jung J H, Domijan M, Klose C, Biswas S, Ezer D, Gao M, Khattak A K, Box M S, Charoensawan V, Cortijo S, Kumar M, Grant A, Locke J C W, Schäfer E, Jaeger K E, Wigge P A. Phytochromes function as thermosensors in Arabidopsis. Science, 2016,354:886-889.
pmid: 27789797 |
| [17] |
Box M S, Huang B E, Domijan M, Jaeger K E, Khattak A K, Yoo S J, Sedivy E L, Jones D M, Hearn T J, Webb A A R, Grant A, Locke J C W, Wigge P A. ELF3 controls thermoresponsive growth in Arabidopsis. Curr Biol, 2015,25:194-199.
doi: 10.1016/j.cub.2014.10.076 pmid: 25557663 |
| [18] |
Tasset C, Singh Yadav A, Sureshkumar S, Singh R, van der Woude L, Nekrasov M, Tremethick D, van Zanten M, Balasubramanian S. POWERDRESS-mediated histone deacetylation is essential for thermomorphogenesis in Arabidopsis thaliana. PLoS Genet, 2018,14:e1007280.
pmid: 29547672 |
| [19] |
Lee C M, Thomashow M F. Photoperiodic regulation of the C-repeat binding factor (CBF) cold acclimation pathway and freezing tolerance in Arabidopsis thaliana. Proc Natl Acad Sci USA, 2012,109:15054-15059.
pmid: 22927419 |
| [20] |
Liu Z J, Zhang Y Q, Wang J F, Li P, Zhao C Z, Chen Y D, Bi Y R. Phytochrome-interacting factors PIF4 and PIF5 negatively regulate anthocyanin biosynthesis under red light in Arabidopsis seedlings. Plant Sci, 2015,238:64-72.
pmid: 26259175 |
| [21] |
Gangappa S N, Berriri S, Kumar S V. PIF4 coordinates thermosensory growth and immunity in arabidopsis. Curr Biol, 2017,27:243-249.
doi: 10.1016/j.cub.2016.11.012 pmid: 28041792 |
| [22] |
Sun J Q, Qi L L, Li Y N, Chu J F, Li C Y. PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet, 2012,8:e1002594.
pmid: 22479194 |
| [23] |
Franklin K A, Lee S H, Patel D, Kumar S V, Spartz A K, Gu C, Ye S Q, Yu P, Breen G, Cohen J D, Wigge P A, Gray W M. Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc Natl Acad Sci USA, 2011,108:20231-20235.
pmid: 22123947 |
| [24] |
Hornitschek P, Kohnen M V, Lorrain S, Rougemont J, Ljung K, Lopez-Vidriero I, Franco-Zorrilla J M, Solano R, Trevisan M, Pradervand S, Xenarios I, Fankhauser C. Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. Plant J, 2012,71:699-711.
pmid: 22536829 |
| [25] | Oh E, Zhu J Y, Bai M Y, Arenhart R A, Sun Y, Wang Z Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. eLife, 2014,3:e03031. |
| [26] |
Boure N, Kumar S V, Arnaud N. The BAP Module: a multisignal integrator orchestrating growth. Trends Plant Sci, 2019,24:602-610.
doi: 10.1016/j.tplants.2019.04.002 pmid: 31076166 |
| [27] |
Min L, Li Y Y, Hu Q, Zhu L F, Gao W H, Wu Y L, Ding Y H, Liu S M, Yang X Y, Zhang X L. Sugar and auxin signaling pathways respond to high-temperature stress during anther development as revealed by transcript profiling analysis in cotton. Plant Physiol, 2014,164:1293-1308.
pmid: 24481135 |
| [28] |
Min L, Zhu L F, Tu L L, Deng F L, Yuan D J, Zhang X L. Cotton GhCKI disrupts normal male reproduction by delaying tapetum programmed cell death via inactivating starch synthase. Plant J, 2013,75:823-835.
doi: 10.1111/tpj.12245 pmid: 23662698 |
| [29] | Min L, Hu Q, Li Y Y, Xu J, Ma Y Z, Zhu L F, Yang X Y, Zhang X L. LEAFY COTYLEDON1-CASEIN KINASE I-TCP15-PHYT OCHROME INTERACTING FACTOR4 network regulates somatic embryogenesis by regulating auxin homeostasis. Plant Physiol, 2015,169:2805-2821. |
| [30] | 胡海燕. 棉花茉莉酸信号负调控因子GhJAZ2的克隆及功能分析. 华中农业大学博士学位论文, 湖北武汉, 2016. |
| Hu H Y. The Cloning and Functional Characterization of a Cotton Negative Regulater GhJAZ2 in Jasmonic Acid Pathway. PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei, China, 2016 (in Chinese with English abstract). | |
| [31] | Jin S X, Zhang X L, Nie Y C, Guo X P, Liang S G, Zhu H G. Identification of a novel elite genotype for in vitro culture and genetic transformation of cotton. Biol Plant, 2006,50:519-524. |
| [32] |
Wu Y L, Min L, Wu Z C, Yang L, Zhu L F, Yang X Y, Yuan D J, Guo X P, Zhang X L. Defective pollen wall contributes to male sterility in the male sterile line 1355A of cotton. Sci Rep, 2015,5:9608.
doi: 10.1038/srep09608 pmid: 26043720 |
| [33] |
Yao X, Tian L, Yang J, Zhao Y N, Zhu Y X, Dai X, Zhao Y, Yang Z N. Auxin production in diploid microsporocytes is necessary and sufficient for early stages of pollen development. PLoS Genet, 2018,14:e1007397.
doi: 10.1371/journal.pgen.1007397 pmid: 29813066 |
| [34] |
Feng X L, Ni W M, Elge S, Mueller-Roeber B, Xu Z H, Xue H W. Auxin flow in anther filaments is critical for pollen grain development through regulating pollen mitosis. Plant Mol Biol, 2006,61:215-226.
doi: 10.1007/s11103-006-0005-z pmid: 16786302 |
| [35] |
Sun J Q, Qi L L, Li Y N, Zhai Q Z, Li C Y. PIF4 and PIF5 transcription factors link blue light and auxin to regulate the phototropic response in Arabidopsis. Plant Cell, 2013,25:2102-2114.
doi: 10.1105/tpc.113.112417 pmid: 23757399 |
| [36] |
Cheng Y F, Dai X H, Zhao Y D. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev, 2006,20:1790-1799.
pmid: 16818609 |
| [37] |
Tadashi S, Takeshi O, Shinya M, Mari T, Yuta T, Nahoko H, Yutaka M, Hideyuki T, Masao W, Atsushi H. Auxins reverse plant male sterility caused by high temperatures. Proc Natl Acad Sci USA, 2010,107:8569-8574.
doi: 10.1073/pnas.1000869107 pmid: 20421476 |
| [38] |
Cecchetti V, Altamura M M, Brunetti P, Petrocelli V, Falasca G, Ljung K, Costantino P, Cardarelli M. Auxin controls Arabidopsis anther dehiscence by regulating endothecium lignification and jasmonic acid biosynthesis. Plant J, 2013,74:411-422.
pmid: 23410518 |
| [39] |
Cecchetti V, Altamura M M, Falasca G, Costantino P, Cardarelli M. Auxin regulates Arabidopsis anther dehiscence, pollen maturation, and filament elongation. Plant Cell, 2008,20:1760-1774.
doi: 10.1105/tpc.107.057570 pmid: 18628351 |
| [40] |
Goldberg R B, Beals T P, Sanders P M. Anther development: basic principles and practical applications. Plant Cell, 1993,5:1217-1229.
doi: 10.1105/tpc.5.10.1217 pmid: 8281038 |
| [41] |
Yang C Y, Xu Z Y, Song J, Conner K, Vizcay Barrena G, Wilson Z A. Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening in the endothecium and is essential for anther dehiscence. Plant Cell, 2007,19:534-548.
doi: 10.1105/tpc.106.046391 pmid: 17329564 |
| [42] |
Stepanova A N, Robertson-Hoyt J, Yun J, Benavente L M, Xie D Y, Dolezal K, Schlereth A, Jurgens G, Alonso J M. TAA1-mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell, 2008,133:177-191.
doi: 10.1016/j.cell.2008.01.047 pmid: 18394997 |
| [43] |
Muller T M, Bottcher C, Morbitzer R, Gotz C C, Lehmann J, Lahaye T, Glawischnig E. TRANSCRIPTION ACTIVATOR- LIKE EFFECTOR NUCLEASE-mediated generation and metabolic analysis of camalexin-deficient cyp71a12 cyp71a13 double knockout lines. Plant Physiol, 2015,168:849-858.
doi: 10.1104/pp.15.00481 pmid: 25953104 |
| [44] |
Nagpal P, Ellis C M, Weber H, Ploense S E, Barkawi L S, Guilfoyle T J, Hagen G, Alonso J M, Cohen J D, Farmer E E, Ecker J R, Reed J W. Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development, 2005,132:4107-4118.
doi: 10.1242/dev.01955 pmid: 16107481 |
| [45] |
Wu M F, Tian Q, Reed J W. Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development, 2006,133:4211-4218.
pmid: 17021043 |
| [1] | 周静远, 孔祥强, 张艳军, 李雪源, 张冬梅, 董合忠. 基于种子萌发出苗过程中弯钩建成和下胚轴生长的棉花出苗壮苗机制与技术[J]. 作物学报, 2022, 48(5): 1051-1058. |
| [2] | 孙思敏, 韩贝, 陈林, 孙伟男, 张献龙, 杨细燕. 棉花苗期根系分型及根系性状的关联分析[J]. 作物学报, 2022, 48(5): 1081-1090. |
| [3] | 闫晓宇, 郭文君, 秦都林, 王双磊, 聂军军, 赵娜, 祁杰, 宋宪亮, 毛丽丽, 孙学振. 滨海盐碱地棉花秸秆还田和深松对棉花干物质积累、养分吸收及产量的影响[J]. 作物学报, 2022, 48(5): 1235-1247. |
| [4] | 郑曙峰, 刘小玲, 王维, 徐道青, 阚画春, 陈敏, 李淑英. 论两熟制棉花绿色化轻简化机械化栽培[J]. 作物学报, 2022, 48(3): 541-552. |
| [5] | 张艳波, 王袁, 冯甘雨, 段慧蓉, 刘海英. 棉籽油分和3种主要脂肪酸含量QTL分析[J]. 作物学报, 2022, 48(2): 380-395. |
| [6] | 张特, 王蜜蜂, 赵强. 滴施缩节胺与氮肥对棉花生长发育及产量的影响[J]. 作物学报, 2022, 48(2): 396-409. |
| [7] | 荐红举, 尚丽娜, 金中辉, 丁艺, 李燕, 王季春, 胡柏耿, Vadim Khassanov, 吕典秋. 马铃薯PIF家族成员鉴定及其对高温胁迫的响应分析[J]. 作物学报, 2022, 48(1): 86-98. |
| [8] | 赵文青, 徐文正, 杨锍琰, 刘玉, 周治国, 王友华. 棉花叶片响应高温的差异与夜间淀粉降解密切相关[J]. 作物学报, 2021, 47(9): 1680-1689. |
| [9] | 岳丹丹, 韩贝, Abid Ullah, 张献龙, 杨细燕. 干旱条件下棉花根际真菌多样性分析[J]. 作物学报, 2021, 47(9): 1806-1815. |
| [10] | 曾紫君, 曾钰, 闫磊, 程锦, 姜存仓. 低硼及高硼胁迫对棉花幼苗生长与脯氨酸代谢的影响[J]. 作物学报, 2021, 47(8): 1616-1623. |
| [11] | 马欢欢, 方启迪, 丁元昊, 池华斌, 张献龙, 闵玲. 棉花GhMADS7基因正调控棉花花瓣发育[J]. 作物学报, 2021, 47(5): 814-826. |
| [12] | 许乃银, 赵素琴, 张芳, 付小琼, 杨晓妮, 乔银桃, 孙世贤. 基于GYT双标图对西北内陆棉区国审棉花品种的分类评价[J]. 作物学报, 2021, 47(4): 660-671. |
| [13] | 周冠彤, 雷建峰, 代培红, 刘超, 李月, 刘晓东. 棉花CRISPR/Cas9基因编辑有效sgRNA高效筛选体系的研究[J]. 作物学报, 2021, 47(3): 427-437. |
| [14] | 卢合全, 唐薇, 罗振, 孔祥强, 李振怀, 徐士振, 辛承松. 商品有机肥替代部分化肥对连作棉田土壤养分、棉花生长发育及产量的影响[J]. 作物学报, 2021, 47(12): 2511-2521. |
| [15] | 王晔, 刘钊, 肖爽, 李芳军, 吴霞, 王保民, 田晓莉. 转PSAG12-IPT基因对棉花叶片衰老及产量和纤维品质的影响[J]. 作物学报, 2021, 47(11): 2111-2120. |
|
||