欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (10): 1628-1638.doi: 10.3724/SP.J.1006.2020.94166
李娜娜1( ), 刘莹1,2, 张豪杰1, 王璐1, 郝心愿1, 张伟富1, 王玉春1, 熊飞1,3, 杨亚军1,*(), 王新超1,*()
), 刘莹1,2, 张豪杰1, 王璐1, 郝心愿1, 张伟富1, 王玉春1, 熊飞1,3, 杨亚军1,*(), 王新超1,*()
LI Na-Na1(), LIU Ying1,2, ZHANG Hao-Jie1, WANG Lu1, HAO Xin-Yuan1, ZHANG Wei-Fu1, WANG Yu-Chun1, XIONG Fei1,3, YANG Ya-Jun1,*(), WANG Xin-Chao1,*()
摘要:
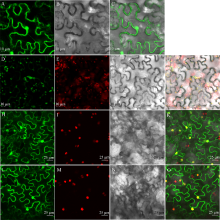
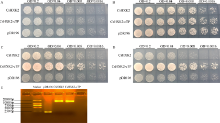
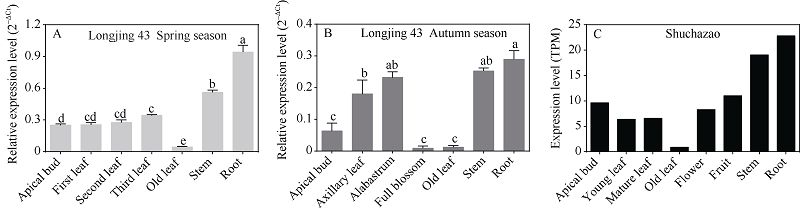
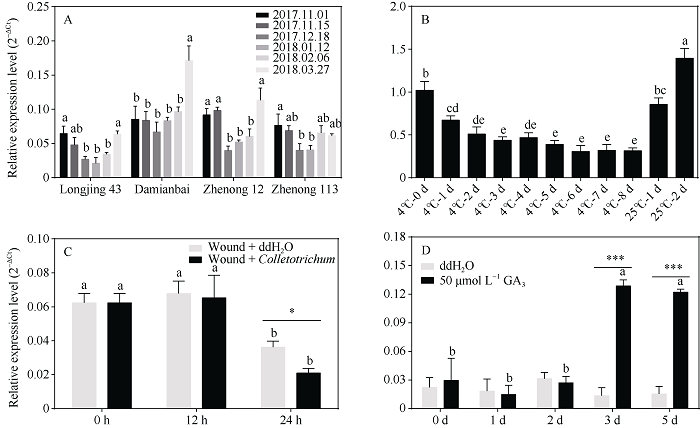
植物己糖激酶是双功能蛋白, 具有磷酸化己糖和介导糖信号的关键性作用。前期研究中, 我们从茶树中克隆获得4个己糖激酶基因, 其中CsHXK2基因编码492个氨基酸残基, 与拟南芥AtHXK3、番茄LeHXK4归为Type A类HXKs。利用RT-PCR技术, 克隆获得长度为2029 bp的CsHXK2基因启动子。CsHXK2基因可能受到光照、低温、病原菌、糖和多种激素等信号的调控, 且可能特异性表达于叶、花、种子、根系、腋芽等组织。CsHXK2蛋白定位于叶绿体内。酵母突变体功能互补试验表明, 去除叶绿体转运信号肽的CsHXK2成熟蛋白具有葡萄糖和果糖磷酸化活性。茶树组织特异性表达分析显示, CsHXK2基因在根和茎中表达量最高, 而在老叶中表达量最低。CsHXK2基因的表达受低温胁迫而显著下调, 经炭疽菌侵染的茶树叶片内CsHXK2基因的表达也受到显著抑制, 而外源赤霉素(GA3)处理的茶树叶片内CsHXK2基因表达显著上调。本研究结果表明, CsHXK2基因在茶树的生长发育过程和逆境胁迫响应中发挥重要的调控作用。
| [1] |
Xiao W Y, Sheen J, Jang J C. The role of hexokinase in plant sugar signal transduction and growth and development. Plant Mol Biol, 2000,44:451-461.
doi: 10.1023/A:1026501430422 |
| [2] |
Rolland F, Sheen J. Sugar sensing and signalling networks in plants. Biochem Soc Trans, 2005,33:269-271.
doi: 10.1042/BST0330269 pmid: 15667323 |
| [3] |
Baena González E, Sheen J. Convergent energy and stress signaling. Trends Plant Sci, 2008,13:474-482.
doi: 10.1016/j.tplants.2008.06.006 pmid: 18701338 |
| [4] |
Dai N, Schaffer A A, Petreikov M, Granot D. Arabidopsis thaliana hexokinase cDNA isolated by complementation of yeast cells. Plant Physiol, 1995,108:879-880.
doi: 10.1104/pp.108.2.879 pmid: 7610198 |
| [5] |
Aguilera Alvarado G P, Sánchez Nieto S. Plant hexokinases are multifaceted proteins. Plant Cell Physiol, 2017,58:1151-1160.
doi: 10.1093/pcp/pcx062 pmid: 28449056 |
| [6] |
Geng M T, Yao Y, Wang Y L, Wu X H, Sun C, Li R M, Fu S P, Duan R J, Liu J, Hu X W, Guo J C. Structure, expression, and functional analysis of the hexokinase gene family in Cassava. Int J Mol Sci, 2017,18:1041.
doi: 10.3390/ijms18051041 |
| [7] |
Wang J X, Wang X M, Geng S Y, Singh S K, Wang Y H, Pattanaik S, Yuan L. Genome-wide identification of hexokinase gene family in Brassica napus: structure, phylogenetic analysis, expression, and functional characterization. Planta, 2018,248:171-182.
doi: 10.1007/s00425-018-2888-8 pmid: 29644447 |
| [8] |
Zhao B Y, Qi K J, Yi X R, Chen G D, Liu X, Qi X X, Zhang S L. Identification of hexokinase family members in pear (Pyrus × bretschneideri) and functional exploration of PbHXK1 in modulating sugar content and plant growth. Gene, 2019,711:143932.
doi: 10.1016/j.gene.2019.06.022 pmid: 31202905 |
| [9] |
Wang H B, Xin H, Guo J Y, Gao Y, Liu C, Dai D Q, Tang L Z. Genome-wide screening of hexokinase gene family and functional elucidation of HXK2 response to cold stress in Jatropha curcas. Mol Biol Rep, 2019,46:1649-1660.
doi: 10.1007/s11033-019-04613-0 pmid: 30756333 |
| [10] |
Karve A, Rauh B L, Xia X X, Kandasamy M, Meagher R B, Sheen J, Moore B D. Expression and evolutionary features of the hexokinase gene family in Arabidopsis. Planta, 2008,228:411-425.
doi: 10.1007/s00425-008-0746-9 |
| [11] |
Kim Y M, Heinzel N, Giese J O, Koeber J, Melzer M, Rutten T, Von Wirén N, Sonnewald U, Hajirezaei M R. A dual role of tobacco hexokinase 1 in primary metabolism and sugar sensing. Plant Cell Environ, 2013,36:1311-1327.
doi: 10.1111/pce.12060 pmid: 23305564 |
| [12] |
Cho J I, Ryoo N, Ko S, Lee S K, Lee J, Jung K H, Lee Y H, Bhoo S H, Winderickx J, An G, Hahn T R, Jeon J S. Structure, expression, and functional analysis of the hexokinase gene family in rice (Oryza sativa L.). Planta, 2006,224:598-611.
doi: 10.1007/s00425-006-0251-y |
| [13] |
Olsson T, Thelander M, Ronne H. A novel type of chloroplast stromal hexokinase is the major glucose-phosphorylating enzyme in the moss Physcomitrella patens. J Biol Chem, 2003,278:44439-44447.
doi: 10.1074/jbc.M306265200 pmid: 12941966 |
| [14] |
Claeyssen É, Rivoal J. Isozymes of plant hexokinase: occurrence, properties and functions. Phytochemistry, 2007,68:709-731.
doi: 10.1016/j.phytochem.2006.12.001 pmid: 17234224 |
| [15] |
Jang J C, León P, Zhou L, Sheen J. Hexokinase as a sugar sensor in higher plants. Plant Cell, 1997,9:5-19.
doi: 10.1105/tpc.9.1.5 pmid: 9014361 |
| [16] |
Dai N, Schaffer A, Petreikov M, Shahak Y, Giller Y, Ratner K, Levine A, Granot D. Overexpression of Arabidopsis hexokinase in tomato plants inhibits growth, reduces photosynthesis, and induces rapid senescence. Plant Cell, 1999,11:1253-1266.
doi: 10.1105/tpc.11.7.1253 pmid: 10402427 |
| [17] |
Moore B, Zhou L, Rolland F, Hall Q, Cheng W H, Liu Y X, Hwang I, Jones T, Sheen J. Role of the Arabidopsis glucose sensor HXK1 in nutrient, light, and hormonal signaling. Science, 2003,300:332-336.
doi: 10.1126/science.1080585 pmid: 12690200 |
| [18] |
Sarowar S, Lee J Y, Ahn E R, Pai H S. A role of hexokinases in plant resistance to oxidative stress and pathogen infection. J Plant Biol, 2008,51:341-346.
doi: 10.1007/BF03036136 |
| [19] |
Li N N, Qian W J, Wang L, Cao H L, Hao X Y, Yang Y J, Wang X C. Isolation and expression features of hexose kinase genes under various abiotic stresses in the tea plant (Camellia sinensis). J Plant Physiol, 2017,209:95-104.
doi: 10.1016/j.jplph.2016.11.007 pmid: 28013175 |
| [20] |
Xia E H, Zhang H B, Sheng J, Li K, Zhang Q J, Kim C, Zhang Y, Liu Y, Zhu T, Li W, Huang H, Tong Y, Nan H, Shi C, Shi C, Jiang J J, Mao S Y, Jiao J Y, Zhang D, Zhao Y, Zhao Y J, Zhang L P, Liu Y L, Liu B Y, Yu Y, Shao S F, Ni D J, Eichler E E, Gao L Z. The tea tree genome provides insights into tea flavor and independent evolution of caffeine biosynthesis. Mol Plant, 2017,10:866-877.
doi: 10.1016/j.molp.2017.04.002 pmid: 28473262 |
| [21] |
Higo K, Ugawa Y, Iwamoto M, Korenaga T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res, 1999,27:297-300.
doi: 10.1093/nar/27.1.297 pmid: 9847208 |
| [22] |
De Winde J H, Crauwels M, Hohmann S, Thevelein J M, Winderickx J. Differential requirement of the yeast sugar kinases for sugar sensing in establishing the catabolite-repressed state. Eur J Biochem, 1996,241:633-643.
doi: 10.1111/j.1432-1033.1996.00633.x pmid: 8917466 |
| [23] |
Hao X Y, Horvath D P, Chao W S, Yang Y J, Wang X C, Xiao B. Identification and evaluation of reliable reference genes for quantitative real-time PCR analysis in tea plant (Camellia sinensis (L.) O. Kuntze). Int J Mol Sci, 2014,15:22155-22172.
doi: 10.3390/ijms151222155 pmid: 25474086 |
| [24] |
Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method . Methods, 2001,25:402-408.
doi: 10.1006/meth.2001.1262 pmid: 11846609 |
| [25] |
Xia E H, Li F D, Tong W, Li P H, Wu Q, Zhao H J, Ge R H, Li R P, Li Y Y, Zhang Z Z, Wei C L, Wan X C. Tea Plant Information Archive: a comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnol J, 2019,17:1938-1953.
doi: 10.1111/pbi.13111 pmid: 30913342 |
| [26] |
Wang L, Yao L N, Hao X Y, Li N N, Wang Y C, Ding C Q, Lei L, Qian W J, Zeng J M, Yang Y J, Wang X C. Transcriptional and physiological analyses reveal the association of ROS metabolism with cold tolerance in tea plant. Environ Exp Bot, 2019,160:45-58.
doi: 10.1016/j.envexpbot.2018.11.011 |
| [27] |
Granot D, David Schwartz R, Kelly G. Hexose kinases and their role in sugar-sensing and plant development. Front Plant Sci, 2013,4:1-17.
doi: 10.3389/fpls.2013.00001 pmid: 23346092 |
| [28] |
Giese J O, Herbers K, Hoffmann M, Klösgen R B, Sonnewald U. Isolation and functional characterization of a novel plastidic hexokinase from Nicotiana tabacum. FEBS Lett, 2005,579:827-831.
doi: 10.1016/j.febslet.2004.12.071 pmid: 15670855 |
| [29] |
Kandel Kfir M, Damari Weissler H, German M A, Gidoni D, Mett A, Belausov E, Petreikov M, Adir N, Granot D. Two newly identified membrane-associated and plastidic tomato HXKs: characteristics, predicted structure and intracellular localization. Planta, 2006,224:1341-1352.
doi: 10.1007/s00425-006-0318-9 |
| [30] |
Yue C, Cao H L, Wang L, Zhou Y H, Huang Y T, Hao X Y, Wang Y C, Wang B, Yang Y J, Wang X C. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season. Plant Mol Biol, 2015,88:591-608.
doi: 10.1007/s11103-015-0345-7 pmid: 26216393 |
| [31] | 田野, 王梦馨, 王金和, 韩宝瑜. 茶鲜叶可溶性糖和氨基酸含量与低温的相关性. 茶叶科学, 2015,35:567-573. |
| Tian Y, Wang M X, Wang J H, Han B Y. Correlation of low temperature with soluble sugar and amino acid content in fresh tea leaves. J Tea Sci, 2015,35:567-573 (in Chinese with English abstract). | |
| [32] |
Lobato A K S, Gonçalves Vidigal M C, Vidigal P S, Costa R C L, Cruz F J R, Santos D G C, Silva C R, Silva L I, Sousa L L. Changes in photosynthetic pigment and carbohydrate content in common bean cultivars infected by Colletotrichum lindemuthianum. Plant Soil Environ, 2009,55:58-61.
doi: 10.17221/PSE |
| [33] |
Engelsdorf T, Horst R J, Pröls R, Pröschel M, Dietz F, Hückelhoven R, Voll L M. Reduced carbohydrate availability enhances the susceptibility of Arabidopsis toward Colletotrichum higginsianum. Plant Physiol, 2013,162:225-238.
doi: 10.1104/pp.112.209676 |
| [34] | Ma Y Y, Zhang Y L, Lu J, Shao H B. Roles of plant soluble sugars and their responses to plant cold stress. Afr J Biotechnol, 2009,8:2004-2010. |
| [35] |
Neta Sharir I, Shoseyov O, Weiss D. Sugars enhance the expression of gibberellin-induced genes in developing petunia flowers. Physiol Plant, 2000,109:196-202.
doi: 10.1034/j.1399-3054.2000.100212.x |
| [36] |
Fukumoto T, Kano A, Ohtani K, Yamasaki Kokudo Y, Kim B G, Hosotani K, Saito M, Shirakawa C, Tajima S, Izumori K, Ohara T, Shigematsu Y, Tanaka K, Ishida Y, Nishizawa Y, Tada Y, Ichimura K, Gomi K, Akimitsu K. Rare sugar D-allose suppresses gibberellin signaling through hexokinase-dependent pathway in Oryza sativa L. Planta, 2011,234:1083-1095.
doi: 10.1007/s00425-011-1463-3 |
| [1] | 冯亚, 朱熙, 罗红玉, 李世贵, 张宁, 司怀军. 马铃薯StMAPK4响应低温胁迫的功能解析[J]. 作物学报, 2022, 48(4): 896-907. |
| [2] | 周悦, 赵志华, 张宏宁, 孔佑宾. 大豆紫色酸性磷酸酶基因GmPAP14启动子克隆与功能分析[J]. 作物学报, 2022, 48(3): 590-596. |
| [3] | 石磊, 苗利娟, 黄冰艳, 高伟, 张忠信, 齐飞艳, 刘娟, 董文召, 张新友. 花生AhFAD2-1基因启动子及5'-UTR内含子功能验证及其低温胁迫应答[J]. 作物学报, 2021, 47(9): 1703-1711. |
| [4] | 王小纯, 王露露, 张志勇, 秦步坛, 于美琴, 韦一昊, 马新明. 小麦谷氨酰胺合成酶同工酶转录特点及其启动子序列分析[J]. 作物学报, 2021, 47(4): 761-769. |
| [5] | 李兰兰, 母丹, 严雪, 杨陆可, 林文雄, 方长旬. OsPAL2;3对水稻化感抑制稗草能力的调控作用[J]. 作物学报, 2021, 47(2): 197-209. |
| [6] | 王珍, 张晓莉, 孟晓静, 姚梦楠, 缪文杰, 袁大双, 朱冬鸣, 曲存民, 卢坤, 李加纳, 梁颖. 甘蓝型油菜丝裂原活化蛋白激酶7基因(BnMAPK7)上游调控因子的鉴定[J]. 作物学报, 2021, 47(12): 2379-2393. |
| [7] | 王珍, 姚梦楠, 张晓莉, 曲存民, 卢坤, 李加纳, 梁颖. 甘蓝型油菜BnMAPK1的原核表达、亚细胞定位及酵母双杂交文库筛选[J]. 作物学报, 2020, 46(9): 1312-1321. |
| [8] | 王丹丹, 柳洪鹃, 王红霞, 张鹏, 史春余. 甘薯蔗糖转运蛋白基因IbSUT3的克隆及功能分析[J]. 作物学报, 2020, 46(7): 1120-1127. |
| [9] | 郑清雷,余陈静,姚坤存,黄宁,阙友雄,凌辉,许莉萍. 甘蔗Rieske Fe/S蛋白前体基因ScPetC的克隆及表达分析[J]. 作物学报, 2020, 46(6): 844-857. |
| [10] | 衡友强,游西龙,王艳. 费尔干猪毛菜病程相关蛋白SfPR1a基因的异源表达增强了烟草对干旱、盐及叶斑病的抗性[J]. 作物学报, 2020, 46(4): 503-512. |
| [11] | 晁毛妮,胡海燕,王润豪,陈煜,付丽娜,刘庆庆,王清连. 陆地棉钾转运体基因GhHAK5启动子的克隆与功能分析[J]. 作物学报, 2020, 46(01): 40-51. |
| [12] | 常建忠,董春林,张正,乔麟轶,杨睿,蒋丹,张彦琴,杨丽莉,吴佳洁,景蕊莲. 小麦抗逆相关基因TaSAP1的5′非翻译区内含子功能分析[J]. 作物学报, 2019, 45(9): 1311-1318. |
| [13] | 张晓红,胡根海,王寒涛,王聪聪,魏恒玲,付远志,喻树迅. 棉花中GhTFL1a和GhTFL1c基因的表达及启动子分析[J]. 作物学报, 2019, 45(3): 469-476. |
| [14] | 王玲,刘峰,戴明剑,孙婷婷,苏炜华,王春风,张旭,毛花英,苏亚春,阙友雄. 甘蔗ScWRKY4基因的克隆与表达特性分析[J]. 作物学报, 2018, 44(9): 1367-1379. |
| [15] | 杨瑞娟,白建荣,闫蕾,苏亮,王秀红,李锐,张丛卓. 玉米低磷胁迫诱导型强启动子P1502-ZmPHR1的克隆与表达分析[J]. 作物学报, 2018, 44(7): 1000-1009. |
|
||