欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (4): 613-625.doi: 10.3724/SP.J.1006.2021.04144
胡冬秀( ), 刘浩(), 洪彦彬, 梁炫强, 陈小平*()
), 刘浩(), 洪彦彬, 梁炫强, 陈小平*()
HU Dong-Xiu(), LIU Hao(), HONG Yan-Bin, LIANG Xuan-Qiang, CHEN Xiao-Ping*()
摘要:
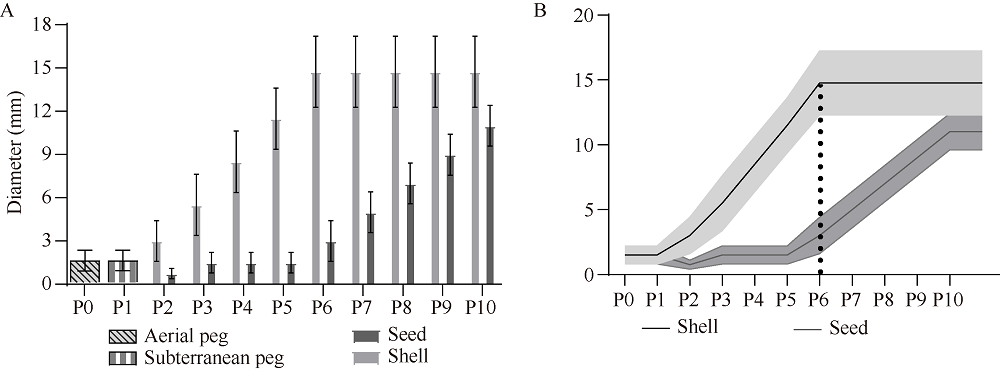
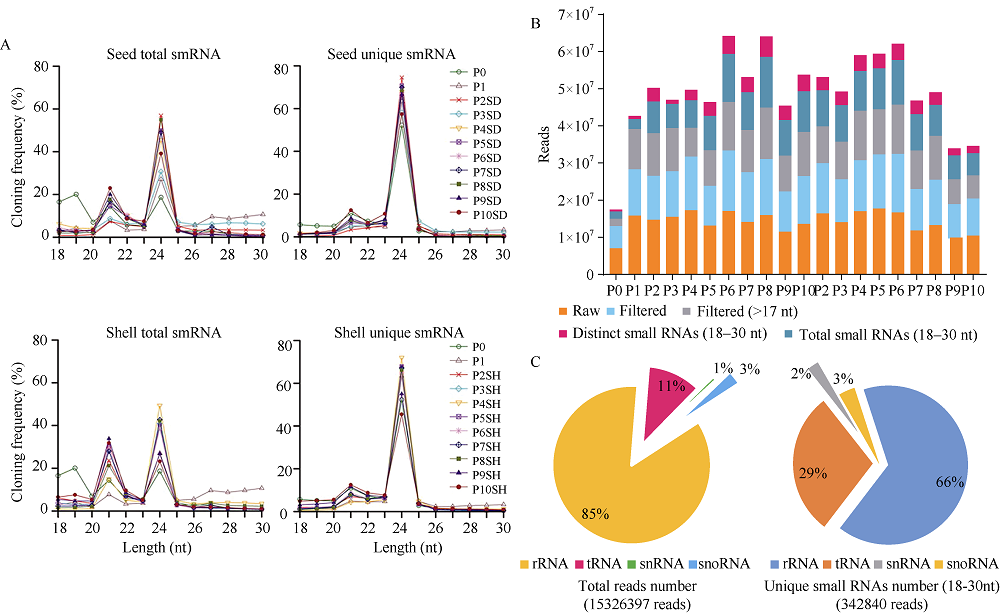
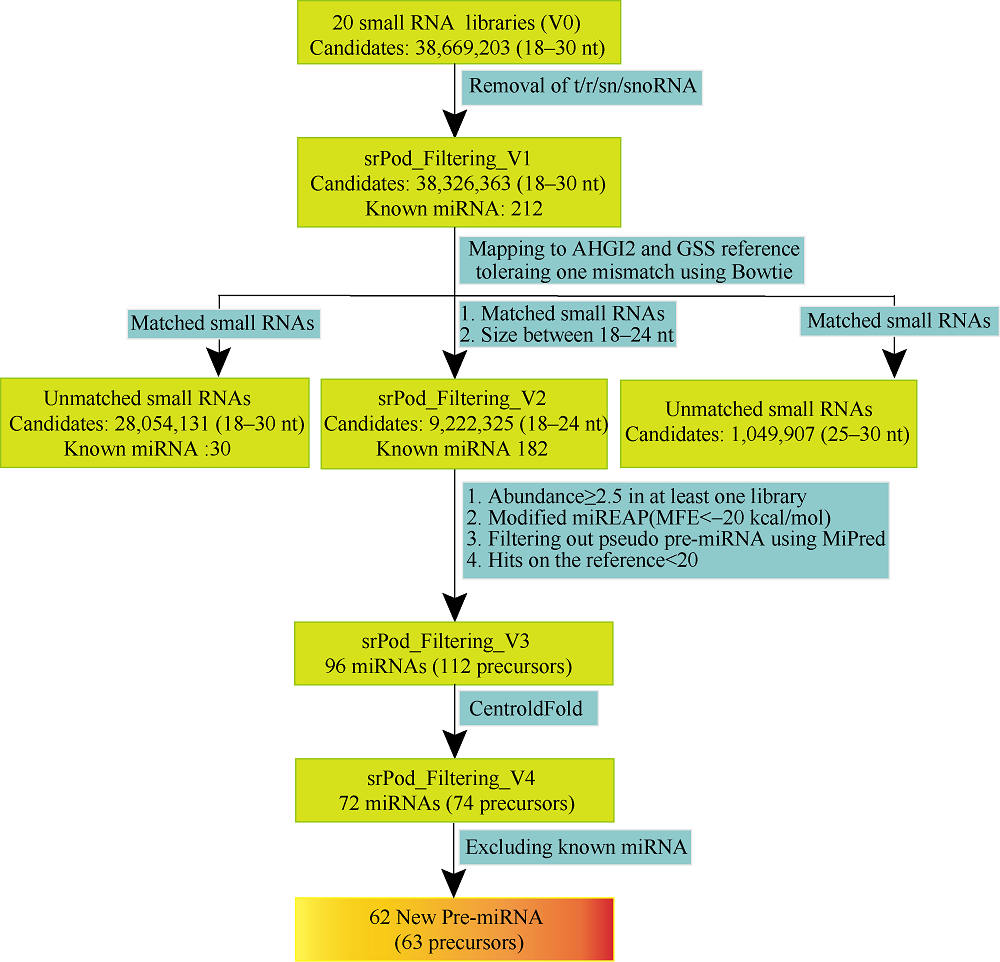
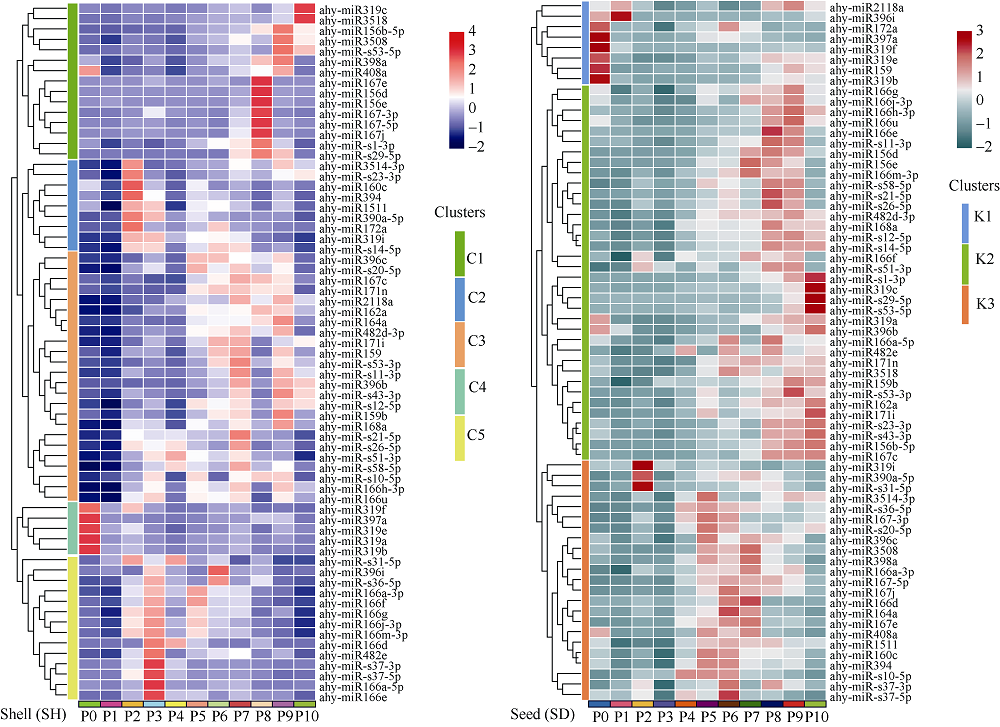
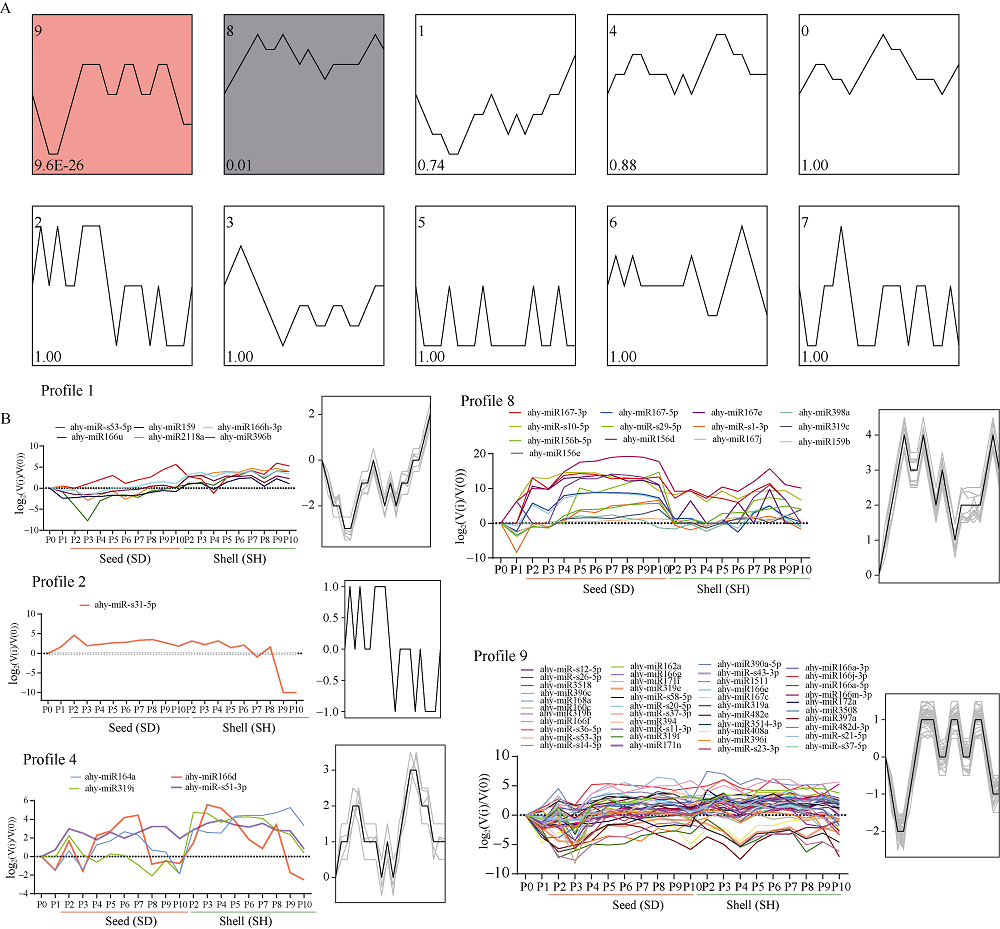
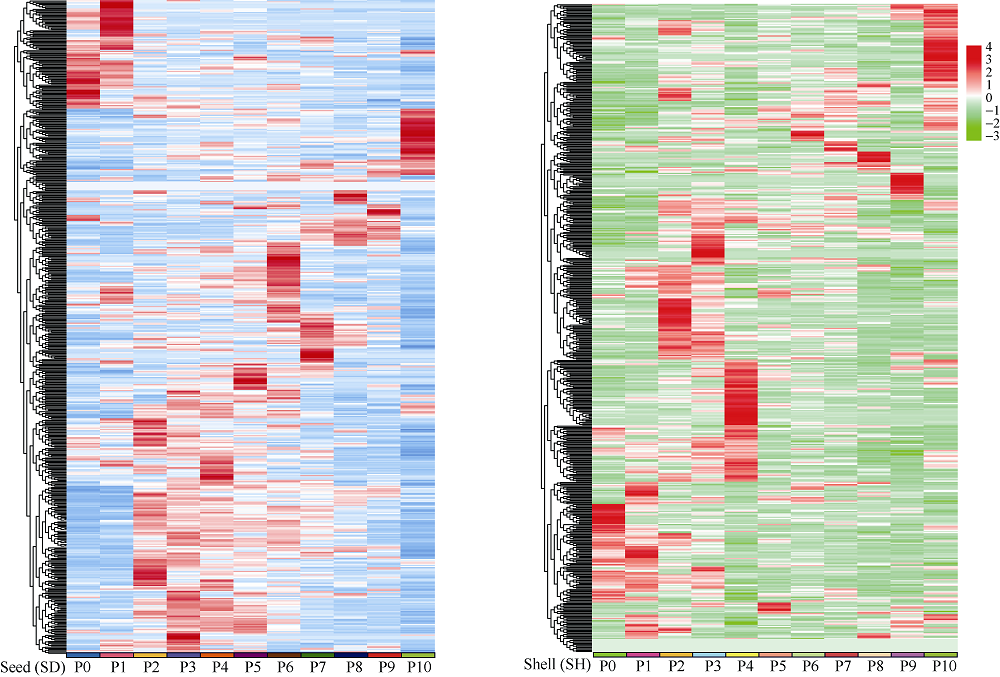
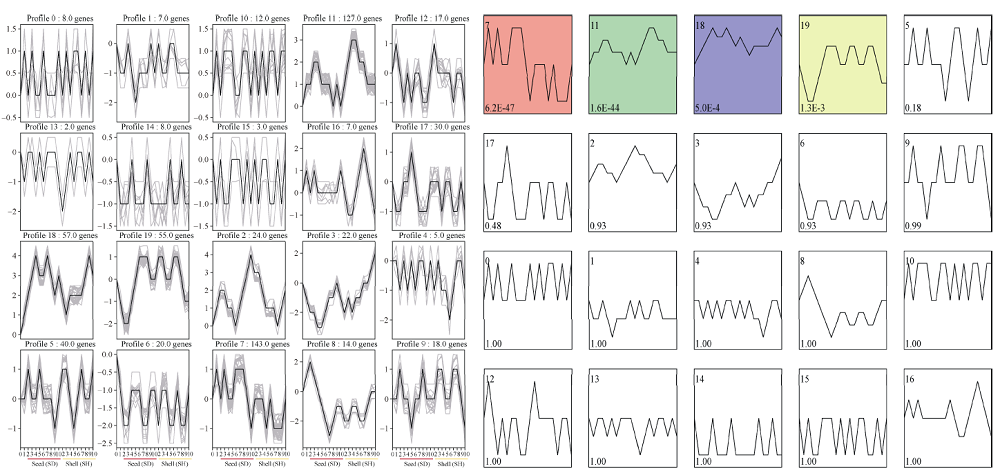
花生(Arachis hypogaea L.)是典型“地上开花、地下结果”的作物, 为从转录后调控水平解析此独特的果实发育现象, 本文应用small RNA测序技术研究荚果发育11个时期果壳及种子中的microRNA及其靶基因。通过测序分别获得212个已知的microRNA和112个新microRNA, 其中, 已知microRNA包括197个保守microRNA和15个花生特异microRNA, 新microRNA来自62个新的microRNA前体序列。表达分析发现, 67个microRNA及其靶基因在荚果发育的11个时期存在时空特异性表达, 部分microRNA的表达量积累阶段性调节果壳与种子的发育, 表明microRNA参与了花生荚果暗发育的整个过程。此外, 对28个microRNA与30个靶基因进行荧光定量PCR验证发现, microRNA和靶基因的表达趋势与测序结果基本一致。本研究通过对花生荚果不同发育时期的果壳和种子进行small RNA测序, 鉴定参与调控花生荚果膨大相关的microRNA, 为研究黑暗条件下植物果实发育的调控机制与花生遗传改良奠定理论基础。
| [1] | Rogers K, Chen X. Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell, 2013,25:2383-2399. |
| [2] |
Chen X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science, 2004,303:2022-2025.
doi: 10.1126/science.1088060 pmid: 12893888 |
| [3] |
Dong Z, Han M H, Fedoroff N. The RNA-binding proteins HYL1 and SE promote accurate in vitro processing of pri-miRNA by DCL1. Proc Natl Acad Sci USA, 2008,105:9970-9975.
pmid: 18632569 |
| [4] |
Yu B, Yang Z Y, Li J J, Minakhina S, Yang M C, Padgett R W, Steward R, Chen X M. Methylation as a crucial step in plant microRNA biogenesis. Science, 2005,307:932-935.
doi: 10.1126/science.1107130 pmid: 15705854 |
| [5] |
Park W, Li J J, Song R T, Messing J, Chen X M. CARPEL FACTORY, a dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana. Curr Biol, 2002,12:1484-1495.
pmid: 12225663 |
| [6] |
Xie M, Zhang S X, Yu B. microRNA biogenesis, degradation and activity in plants. Cell Mol Life Sci, 2015,72:87-99.
doi: 10.1007/s00018-014-1728-7 pmid: 25209320 |
| [7] | 邢利娟, 刘悦萍, 王磊, 徐妙云. miRNA参与植物胚和胚乳发育调控的研究进展. 生物技术进展, 2020,10(2):109-116. |
| Xing L J, Liu Y P, Wang L, Xu M Y. Research progress of miRNA involved in regulation of plant embryo and endosperm development. Curr Biotechnol, 2020,10(2):109-116 (in Chinese with English abstract). | |
| [8] | 秦耀旭, 张关元, 刘司奇, 刘洋, 许志茹. 植物重金属胁迫相关miRNA的研究进展. 分子植物育种, 2019,17:2855-2861. |
| Qin Y X, Zhang G Y, Liu S Q, Liu Y, Xu Z R. Research progress of miRNA related to heavy metal stress in plants. Mol Plant Breed, 2019,17:2855-2861 (in Chinese with English abstract). | |
| [9] | 郁佳雯, 韩荣鹏, 仇婷, 李宗芸, 潘沈元, 董婷婷. microRNA在植物生长发育中的研究进展. 分子植物育种, 2020,18:1496-1504. |
| Yu J W, Han R P, Qiu T, Li Z Y, Pan S Y, Dong T T. Research progress on microRNA in plant growth and development. Mol Plant Breed, 2020,18:1496-1504 (in Chinese with English abstract). | |
| [10] | Long R C, Li M N, Li X, Gao Y L, Zhang T J, Sun Y, Kang J M, Wang T H, Cong L L, Yang Q C. A novel miRNA sponge form efficiently inhibits the activity of miR393 and enhances the salt tolerance and ABA insensitivity in Arabidopsis thaliana. Plant Mol Biol Rep, 2017,35:409-415. |
| [11] | 李科学, 曲德杰, 黄慧梅, 曹金山, 樊仔慧, 李国纪, 王幼宁. miRNA调控大豆根系结瘤及共生固氮的分子机制研究进展. 植物生理学报, 2019,55:1587-1594. |
| Li K X, Qu D J, Huang H M, Cao J S, Fan Z H, Li G J, Wang Y N. Research progress on miRNA-mediated molecular mechanisms of nodulation and symbiotic nitrogen fixation in soybean. J Plant Physiol, 2019,55:1587-1594 (in Chinese with English abstract). | |
| [12] |
Wang T Y, Ping X K, Cao Y R, Jian H J, Gao Y M, Wang J, Tan Y C, Xu X F, Lu K, Li J N, Liu L Z. Genome-wide exploration and characterization of miR172/euAP2 genes in Brassica napus L. for likely role in flower organ development. BMC Plant Biol, 2019,19:336.
doi: 10.1186/s12870-019-1936-2 pmid: 31370790 |
| [13] |
Jiao X M, Wang H C, Yan J J, Kong X Y, Liu Y W, Chu J F, Chen X Y, Fang R X, Yan Y S. Promotion of BR biosynthesis by miR444 is required for ammonium-triggered inhibition of root growth. Plant Physiol, 2020,182:1454-1466.
pmid: 31871071 |
| [14] |
Guo G H, Liu X Y, Sun F L, Cao J, Huo N, Wuda B, Xin M M, Hu Z R, Du J K, Xia R, Rossi V, Peng H R, Ni Z F, Sun Q X, Yao Y Y. Wheat miR9678 affects seed germination by generating phased siRNAs and modulating abscisic acid/gibberellin signaling. Plant Cell, 2018,30:796-814.
pmid: 29567662 |
| [15] |
José Ripoll J, Bailey L J, Mai Q A, Wu S L, Hon C T, Chapman E J, Ditta G S, Estelle M. Yanofsky M F. microRNA regulation of fruit growth. Nat Plants, 2015,1:15036.
doi: 10.1038/nplants.2015.36 pmid: 27247036 |
| [16] |
Todesco M, Rubio-Somoza I, Paz-Ares J, Weigel D. A collection of target mimics for comprehensive analysis of microRNA function in Arabidopsis thaliana. PLoS Genet, 2010,6:e1001031.
doi: 10.1371/journal.pgen.1001031 pmid: 20661442 |
| [17] |
Lu Y Z, Feng Z, Meng Y L, Bian L Y, Xie H, Mysore K S, Liang J S. SLENDER RICE1 and Oryza sativa INDETERMINATE DOMAIN2 regulating osmiR396 are involved in stem elongation. Plant Physiol, 2020,182:2213-2227.
pmid: 31953375 |
| [18] |
Zhang J P, Yu Y, Feng Y Z, Zhou Y F, Zhang F, Yang Y W, Lei M Q, Zhang Y C, Chen Y Q. MiR408 regulates grain yield and photosynthesis via a phytocyanin protein. Plant Physiol, 2017,175:1175-1185.
doi: 10.1104/pp.17.01169 pmid: 28904074 |
| [19] |
Zhao C, Li T, Zhao Y, Zhang B, Li A, Zhao S, Hou L, Xia H, Fan S, Qiu J, Li P, Zhang Y, Guo B, Wang X. Integrated small RNA and mRNA expression profiles reveal miRNAs and their target genes in response to Aspergillus flavus growth in peanut seeds. BMC Plant Biol, 2020,20:215.
pmid: 32404101 |
| [20] |
Ma X L, Zhang X G, Zhao K K, Li F P, Li K, Ning L L, He J L, Xin Z Y, Yin D M. Small RNA and degradome deep sequencing reveals the roles of microRNAs in seed expansion in peanut ( Arachis hypogaea L.). Front Plant Sci, 2018,9:349.
doi: 10.3389/fpls.2018.00349 pmid: 29662498 |
| [21] |
Chen H, Yang Q, Chen K, Zhao S S, Zhang C, Pan R L, Cai T C, Deng Y, Wang X J, Chen Y T, Chu W T, Xie W P, Zhuang W J. Integrated microRNA and transcriptome profiling reveals a miRNA-mediated regulatory network of embryo abortion under calcium deficiency in peanut ( Arachis hypogaea L.). BMC Genomics, 2019,20:392.
pmid: 31113378 |
| [22] |
Gao C, Wang P F, Zhao S Z, Zhao C Z, Xia H, Hou L, Ju Z, Zhang Y, Li C S, Wang X J. Small RNA profiling and degradome analysis reveal regulation of microRNA in peanut embryogenesis and early pod development. BMC Genomics, 2017,18:220.
pmid: 28253861 |
| [23] | Chen C J, Chen H, Zhang Y, Thomas H R, Frank M H, He Y H, Xia R. Tbtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant, 2020,S1674-2052:30187-30188. |
| [24] |
Zhao Y T, Wang M, Fu S X, Yang W C, Qi C K, Wang X J, X J. Small RNA profiling in two Brassica napus cultivars identifies microRNAs with oil production-and development-correlated expression and new small RNA classes. Plant Physiol, 2012,158:813-823.
doi: 10.1104/pp.111.187666 pmid: 22138974 |
| [25] |
Nodine M D, Bartel D P. MicroRNAs prevent precocious gene expression and enable pattern formation during plant embryogenesis. Genes Dev, 2010,24:2678-2692.
pmid: 21123653 |
| [26] |
Palatnik J F, Allen E, Wu X, Schommer C, Schwab R, Carrington J C, Weigel D. Control of leaf morphogenesis by microRNAs. Nature, 2003,425:257-263.
pmid: 12931144 |
| [27] |
Wang S K, Wu K, Yuan Q B, Liu X Y, Liu Z B, Lin X Y, Zeng R Z, Zhu H T, Dong G J, Qian Q, Zhang G Q, Fu X D. Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet, 2012,44:950-954.
doi: 10.1038/ng.2327 pmid: 22729225 |
| [28] |
Pignocchi C, Kiddle G, Hernández I, Foster S J, Asensi A, Taybi T, Barnes J, Foyer C H. Ascorbate oxidase-dependent changes in the redox state of the apoplast modulate gene transcript accumulation leading to modified hormone signaling and orchestration of defense processes in tobacco. Plant Physiol, 2006,141:423-435.
doi: 10.1104/pp.106.078469 pmid: 16603663 |
| [29] |
Potters G, Horemans N, Caubergs R J, Asard H. Ascorbate and dehydroascorbate influence cell cycle progression in a tobacco cell suspension. Plant Physiol, 2000,124:17-20.
pmid: 10982417 |
| [30] |
Yang J H, Han S J, Yoon E K, Lee W S. Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells. Nucleic Acids Res, 2006,34:1892-1899.
doi: 10.1093/nar/gkl118 pmid: 16598073 |
| [31] |
Jones-Rhoades M W, Bartel D P. Computational identification of plant microRNAs and their targets, including a stress induced miRNA. Mol Cell, 2004,14:787-799.
doi: 10.1016/j.molcel.2004.05.027 pmid: 15200956 |
| [1] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [2] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [3] | 刘嘉欣, 兰玉, 徐倩玉, 李红叶, 周新宇, 赵璇, 甘毅, 刘宏波, 郑月萍, 詹仪花, 张刚, 郑志富. 耐三唑并嘧啶类除草剂花生种质创制与鉴定[J]. 作物学报, 2022, 48(4): 1027-1034. |
| [4] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [5] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [6] | 汪颖, 高芳, 刘兆新, 赵继浩, 赖华江, 潘小怡, 毕晨, 李向东, 杨东清. 利用WGCNA鉴定花生主茎生长基因共表达模块[J]. 作物学报, 2021, 47(9): 1639-1653. |
| [7] | 王建国, 张佳蕾, 郭峰, 唐朝辉, 杨莎, 彭振英, 孟静静, 崔利, 李新国, 万书波. 钙与氮肥互作对花生干物质和氮素积累分配及产量的影响[J]. 作物学报, 2021, 47(9): 1666-1679. |
| [8] | 石磊, 苗利娟, 黄冰艳, 高伟, 张忠信, 齐飞艳, 刘娟, 董文召, 张新友. 花生AhFAD2-1基因启动子及5'-UTR内含子功能验证及其低温胁迫应答[J]. 作物学报, 2021, 47(9): 1703-1711. |
| [9] | 高芳, 刘兆新, 赵继浩, 汪颖, 潘小怡, 赖华江, 李向东, 杨东清. 北方主栽花生品种的源库特征及其分类[J]. 作物学报, 2021, 47(9): 1712-1723. |
| [10] | 张鹤, 蒋春姬, 殷冬梅, 董佳乐, 任婧瑶, 赵新华, 钟超, 王晓光, 于海秋. 花生耐冷综合评价体系构建及耐冷种质筛选[J]. 作物学报, 2021, 47(9): 1753-1767. |
| [11] | 薛晓梦, 吴洁, 王欣, 白冬梅, 胡美玲, 晏立英, 陈玉宁, 康彦平, 王志慧, 淮东欣, 雷永, 廖伯寿. 低温胁迫对普通和高油酸花生种子萌发的影响[J]. 作物学报, 2021, 47(9): 1768-1778. |
| [12] | 郝西, 崔亚男, 张俊, 刘娟, 臧秀旺, 高伟, 刘兵, 董文召, 汤丰收. 过氧化氢浸种对花生种子发芽及生理代谢的影响[J]. 作物学报, 2021, 47(9): 1834-1840. |
| [13] | 张旺, 冼俊霖, 孙超, 王春明, 石丽, 于为常. CRISPR/Cas9编辑花生FAD2基因研究[J]. 作物学报, 2021, 47(8): 1481-1490. |
| [14] | 戴良香, 徐扬, 张冠初, 史晓龙, 秦斐斐, 丁红, 张智猛. 花生根际土壤细菌群落多样性对盐胁迫的响应[J]. 作物学报, 2021, 47(8): 1581-1592. |
| [15] | 黄冰艳, 孙子淇, 刘华, 房元瑾, 石磊, 苗利娟, 张毛宁, 张忠信, 徐静, 张梦圆, 董文召, 张新友. 花生巢式群体的脂肪含量遗传分析[J]. 作物学报, 2021, 47(6): 1100-1108. |
|
||