Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2021, Vol. 47 ›› Issue (3): 385-393.doi: 10.3724/SP.J.1006.2021.04123
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Next Articles
YANG Yang( ), LI Huai-Lin(), HU Li-Min, FAN Chu-Chuan*(), ZHOU Yong-Ming
), LI Huai-Lin(), HU Li-Min, FAN Chu-Chuan*(), ZHOU Yong-Ming
| [1] | Mohammad A, Irka S, Aziz M. Inheritance of seed colour in some Brassica oleiferous. Indian J Genet Plant Breed, 1942,2:112-127. |
| [2] | 何余堂, 龙卫华, 胡进平, 傅廷栋, 李殿荣, 陈宝元, 涂金星. 白菜型油菜角果多室性状的遗传及解剖学研究. 中国油料作物学报, 2003,25:1-4. |
| He Y T, Long W H, Hu J P, Fu T D, Li D R, Chen B Y, Tu J X. Anatomic and genetic studies on multicapsular character in Brassica campestris L. Chin J Oil Crop Sci, 2003,25:1-4 (in Chinese with English abstract). | |
| [3] | 刘后利. 油菜的遗传育种学, 北京: 中国农业大学出版社, 2000. p 162. |
| Liu H L. Genetics and Breeding of Brassica napus. Beijing: China Agricultural University Press, 2000. p 162 (in Chinese). | |
| [4] | Varshney S. Inheritance of siliqua characters in Indian colza: I. Locule number and siliqua position. Euphytica, 1987,36:541-544. |
| [5] | 赵洪朝. 芥菜型油菜多室性状的遗传表现及其与甘蓝型油菜的种间远缘杂交. 西北农林科技大学硕士学位论文, 陕西杨凌, 2003. |
| Zhao H C. Study on Multilocular Heredity of Brassica juncea and Interspecific Hybridization between B. juncea and B. napus. MS Thesis of Northwest A&F University, Yangling, Shaanxi, China, 2003 (in Chinese with English abstract). | |
| [6] | 吕泽文. 芥菜型油菜多室性状的遗传分析及其基因的分子标记. 华中农业大学硕士学位论文, 湖北武汉, 2011. |
| Lyu Z W. Study on Inheritance and Molecular Markers for Multilocular Trait in Brassica juncea. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2011 (in Chinese with English abstract). | |
| [7] | Kadkol G P, Halloran G M. Inheritance of siliqua strength in Brassica campestris L.: I. Studies of F2 and backcross populations. Can J Gene Cytol, 1986,28:365-373. |
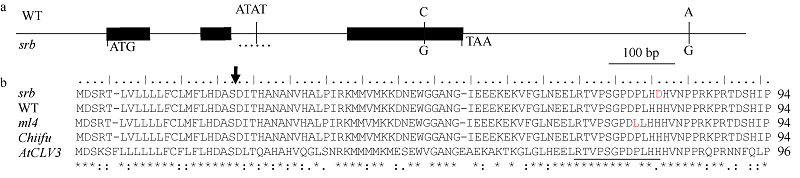
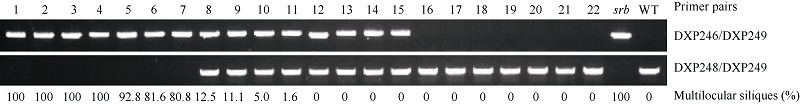
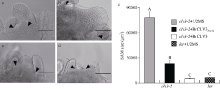
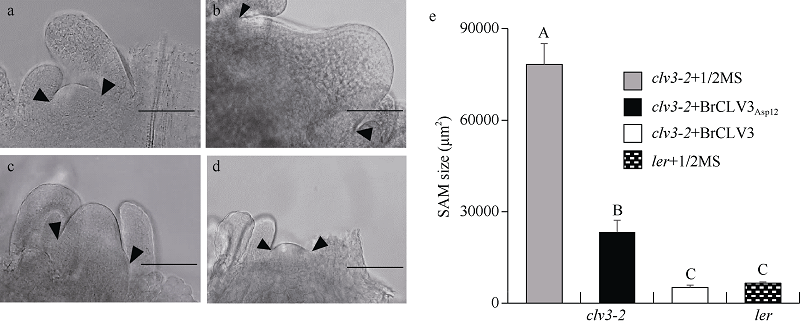
| [8] | Fan C C, Wu Y D, Yang Q Y, Yang Y, Meng Q W, Zhang K Q, Li J G, Wang J F, Zhou Y M. A novel single-nucleotide mutation in a CLAVATA3 gene homologue controls a multilocular silique trait in Brassica rapa L. Mol Plant, 2014,7:1788-1792. |
| [9] | Choudhary B, Solanki Z. Inheritance of siliqua locule number and seed coat colour in Brassica juncea. Plant Breed, 2007,126:104-106. |
| [10] | Xiao L, Zhao H Y, Zhao Z, Du D Z, Xu L, Yao Y M, Zhao Z G, Xing X R, Shang G X, Zhao H C. Genetic and physical fine mapping of a multilocular geneBjln1 in Brassica juncea to a 208-kb region. Mol Breed, 2013,32:373-383. |
| [11] | Xu P, Lyu Z W, Zhang X X, Wang X H, Pu Y Y, Wang H M, Yi B, Wen J, Ma C Z, Tu J X, Fu T D, Shen J X. Identification of molecular markers linked to trilocular gene (mc1) in Brassica juncea L. Mol Breed, 2014,33:425-434. |
| [12] |
Xiao L, Li X, Liu F, Zhao Z, Xu L, Chen C P, Wang Y H, Shang G X, Du D Z. Mutations in the CDS and promoter of BjuA07.CLV1 cause a multilocular trait in Brassica juncea. Sci Rep, 2018,8:5339.
doi: 10.1038/s41598-018-23636-4 pmid: 29593311 |
| [13] | Brand U, Fletcher J C, Hobe M, Meyerowitz E M, Simon R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science, 2000,289:617-619. |
| [14] |
Schoof H, Lenhard M, Haecker A, Mayer K F, Jürgens G, Laux T. The stem cell population ofArabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell, 2000,100:635-644.
pmid: 10761929 |
| [15] |
Yang Y, Zhu K Y, Li H L, Han S Q, Meng Q W, Khan S U, Fan C C, Xie K B, Zhou Y M. Precise editing of CLAVATA genes in Brassica napus L. regulates multilocular silique development. Plant Biotechnol J, 2018,16:1322-1335.
doi: 10.1111/pbi.12872 pmid: 29250878 |
| [16] |
Drenkard E, Richter B, Rozen S, Stutius L M, Angell N A, Mindrinos M, Cho R J, Oefner P J, Davis R W, Ausubel F M. A simple procedure for the analysis of single nucleotide polymorphisms facilitates map-based cloning in Arabidopsis. Plant Physiol, 2000,124:1483-1492.
doi: 10.1104/pp.124.4.1483 pmid: 11115864 |
| [17] |
Kondo T, Nakamura T, Yokomine K, Sakagami Y. Dual assay for MCLV3 activity reveals structure-activity relationship of CLE peptides. Biochem Biophys Res Commun, 2008,377:312-316.
doi: 10.1016/j.bbrc.2008.09.139 |
| [18] |
Song X F, Yu D L, Xu T T, Ren S C, Guo P, Liu C M. Contributions of individual amino acid residues to the endogenous CLV3 function in shoot apical meristem maintenance in Arabidopsis. Mol Plant, 2012,5:515-523.
doi: 10.1093/mp/ssr120 |
| [19] |
Fiers M, Golemiec E, van der Schors R, van der Geest L, Li K W, Stiekema W J, Liu C M. TheCLV3/ESR motif of CLV3 is functionally independent from the nonconserved flanking sequences. Plant Physiol, 2006,141:1284-1292.
pmid: 16751438 |
| [20] |
Kondo T, Sawa S, Kinoshita A, Mizuno S, Kakimoto T, Fukuda H, Sakagami Y. A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science, 2006,313:845-848.
pmid: 16902141 |
| [1] | SU Ya-Chun, LI Cong-Na, SU Wei-Hua, YOU Chui-Huai, CEN Guang-Li, ZHANG Chang, REN Yong-Juan, QUE You-Xiong. Identification of thaumatin-like protein family in Saccharum spontaneum and functional analysis of its homologous gene in sugarcane cultivar [J]. Acta Agronomica Sinica, 2021, 47(7): 1275-1296. |
| [2] | Li-Ping QIN,Er-Fei DONG,Yang BAI,Lian ZHOU,Lan-Yang REN,Ren-Feng ZHANG,Chao-Xian LIU,Yi-Lin CAI. Genetic analysis and molecular characterization of tasselseed mutant ts12 in maize [J]. Acta Agronomica Sinica, 2020, 46(5): 690-699. |
| [3] | Zhi-Fei XUE, Xia WANG, Fu-Peng LI, Chao-Zhi MA. Homologous Cloning of BnGS3 and BnGhd7 Genes in Brassica napus and Their Relationship with Rapeseed Yield-related Traits [J]. Acta Agronomica Sinica, 2018, 44(02): 297-305. |
| [4] | YU Jian,LIU Chang-Ying,ZHAO Ai-Chun,WANG Chuan-Hong,CAI Yu-Xiang,YU Mao-De*. Functional Analysis of 1-Aminocyclopropane-1-carboxylate Oxidase Gene’s Promoter in Mulberry [J]. Acta Agron Sin, 2017, 43(06): 839-848. |
| [5] | YANG Fan,CHEN Qi-Jiao,GAO Xiang,ZHAO Wan-Chun,JIANG Qin-Qin,WU Dan,MENG Min. Cloning, Prokaryotic Expression and in vitro Functional Analysis of α-Gliadin Genes from Dasypyrum villosum [J]. Acta Agron Sin, 2014, 40(08): 1340-1349. |
| [6] | XU Wen-Ting,WANG Cheng,XU Xiao-Yang,NIU Er-Li,CAI Cai-Ping,GUO Wang-Zhen. Cloning and Functional Analysis of GhVacInc2a Encoding Vacuolar Invertase in Cotton [J]. Acta Agron Sin, 2014, 40(03): 390-396. |
| [7] | WANG Rui,WU Hua-Ling,WANG Hui-Fang,HUANG Ke,HUO Chun-Yan,NI Zhong-Fu,SUN Qi-Xin. Cloning, Characterization, and Functional Analysis of TaWRKY44 Gene from Wheat [J]. Acta Agron Sin, 2013, 39(11): 1944-1951. |
| [8] | SUN Yong-Wei, NIE Li-Na, MA Wei-Zhi, XU Zhao-Shi, JIA Lan-Qin. Cloning and Functional Analysis of Viviparous-1 Promoter in Wheat [J]. Acta Agron Sin, 2011, 37(10): 1743-1751. |
|
||