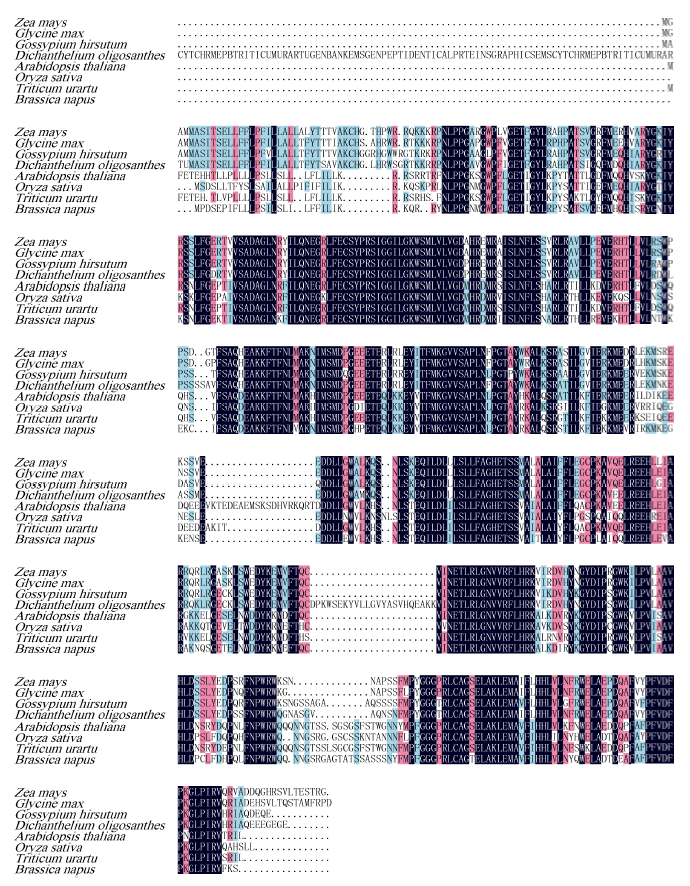
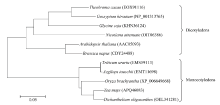
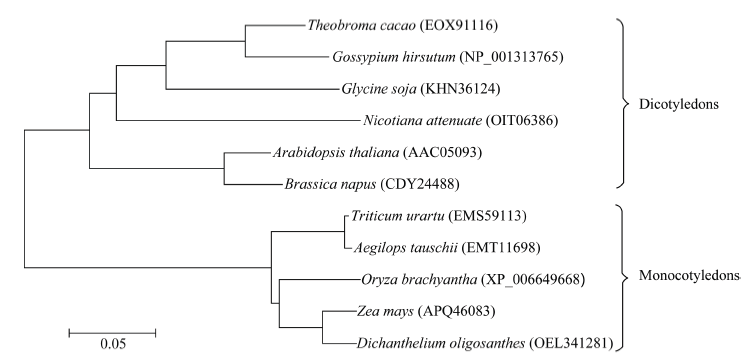
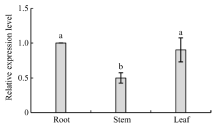
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2018, Vol. 44 ›› Issue (03): 343-356.doi: 10.3724/SP.J.1006.2018.00343
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
Fang-Meng DUAN1( ), Qiu-Lan LUO2, Xue-Li LU3, Na-Wei QI1, Xian-Shun LIU1, Wen-Wen SONG1,*()
), Qiu-Lan LUO2, Xue-Li LU3, Na-Wei QI1, Xian-Shun LIU1, Wen-Wen SONG1,*()
| [1] | Arteca J M, Arteca R N.Brassinosteroid-induced exaggerated growth in hydroponically grown Arabidopsis plants. Physiol Plant, 2001, 112: 104-112 |
| [2] | Hu Y X, Bao F, Li J Y.Promotive effect of brassinosteroids on cell division involves a distinct CycD3-induction pathway in Arabidopsis. Plant J, 2000, 24: 693-701 |
| [3] | Yu J Q, Huang L F, Hu W H, Zhou Y H, Mao W H, Nogués S.A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J Exp Bot, 2004, 55: 1135-1143 |
| [4] | Divi U K, Krishna P.Brassinosteroid: a biotechnological target for enhancing crop yield and stress tolerance.New Biotechnol, 2009, 26: 131-136 |
| [5] | Kurepin L V, Qaderi M M, Back T G, Reid D M, Pharis R P.A rapid effect of applied brassinolide on abscisic acid concentrations in Brassica napus leaf tissue subjected to short-term heat stress. Plant Growth Regul, 2008, 55: 165-167 |
| [6] | El-Mashad A A A, Mohamed H I. Brassinolide alleviates salt stress and increases antioxidant activity of cowpea plants (Vigna sinensis). Protoplasma, 2012, 249: 625-635 |
| [7] | Nakashitaetal H, Yasuda M, Nitta T, Asami T, Fujioka S, Arai Y, Sekimata K, Takatsuto S, Yamaguchi I, Yoshida S.Brassinosteroid function in a broad range of disease resistance in tobacco and rice.Plant J, 2003, 33: 887-898 |
| [8] | 李艳楠. 油菜素内酯处理对番茄根结线虫的防治效果研究. 河南科技大学硕士学位论文, 河南洛阳, 2012 |
| Li Y N.The Research of Control Effects of Brassinosteroids Treatment on Tomato Root-knot Nematode Caused by Meloidogyne Incognita. MS Thesis of Henan University of Science and Technology, Luoyang,China, 2012 (in Chinese with English abstract) | |
| [9] | 张丹丹. 油菜素内酯提高黄瓜对根结线虫抗性的机制研究及田间效应. 浙江大学硕士学位论文, 浙江杭州, 2012 |
| Zhang D D.Field Effect and Molecular Mechanisms of Brassinosteroids-increased the Resistance to Root-knot Nematode in Cucumis sativus. MS Thesis of Zhejiang University, Hangzhou,China, 2012 (in Chinese with English abstract) | |
| [10] | 孙超, 黎家. 油菜素甾醇类激素的生物合成、代谢及信号转导. 植物生理学报, 2017, 53: 291-307 |
| Sun C, Li J.Biosynthesis, catabolism, and signal transduction of brassinosteroids.J Plant Physiol, 2017, 53: 291-307 (in Chinese with English abstract) | |
| [11] | Sahni S, Prasad B D, Liu Q, Grbic V, Sharpe A, Singh S P, Krishnaa P.Overexpression of the brassinosteroid biosynthetic gene DWF4 in Brassica napus simultaneously increases seed yield and stress tolerance. Sci Rep, 2016, 6: 28298 |
| [12] | Shimada Y, Fujioka S, Miyauchi N, Kushiro M, Takatsuto S, Nomura T, Yokota T, Kamiya Y, Bishop G J, Yoshida S.Brassinosteroid-6-oxidases from Arabidopsis and tomato catalyze multiple C-6 oxidations in brassinosteroid biosynthesis. Plant Physiol, 2001, 126: 770-779 |
| [13] | 申强. 过量表达ZmDWF4基因改良玉米产量性状的研究. 山东农业大学硕士学位论文,山东泰安, 2011 |
| Shen Q.Overexpression of ZmDWF4 Improves Yield Traits in Maize. MS Thesis of Shandong Agricultural University, Tai’an,China, 2011 (in Chinese with English abstract) | |
| [14] | Choe S, Fujioka S, Noguchi T, Takatsuto S, Yoshida S, Feldmann K A.Overexpression of DWARF4 in the brassinosteroid biosynthetic pathway results in increased vegetative growth and seed yield in Arabidopsis. Plant J, 2001, 26: 573-582 |
| [15] | 武海军, 司建萍, 王新宇. 胡杨DWF4基因的克隆及其在拟南芥中的异源表达和功能分析. 见: 中国细胞生物学学会主编. 中国细胞生物学学会2013年全国学术大会论文摘要集. 湖北武汉: 2013. p 247 |
| Wu H J, Si J P, Wang X Y.Cloning of the DWF4 gene from Populus euphratica, its heterologous expression and functional analysis in Arabidopsis. In: Chinese Academy of Cell Biology, eds. Proceedings of Academic Conference of Chinese Academy of Cell Biology in 2013. Wuhan, China, 2013. p 247 (in Chinese) | |
| [16] | Wu C Y, Trieu A, Radhakrishnan P, Kwok S F, Harris S, Zhang K, Wang J L, Wan J M, Zhai H Q, Takatsuto S, Matsumoto S, Fujioka S, Feldmann K A, Pennella R I.Brassinosteroids regulate grain filling in rice.Plant Cell, 2008, 20: 2130-2145 |
| [17] | Maharjan P M, Schulz B, Choe S.BIN2/DWF12 antagonistically transduces brassinosteroid and auxin signals in the roots of Arabidopsis. J Plant Physiol, 2011, 54: 126-134 |
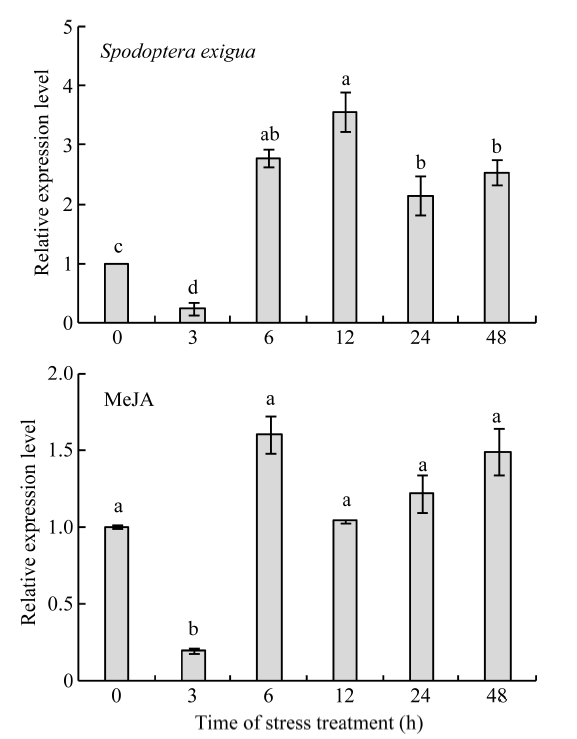
| [18] | 徐涛, 王建斌, 洛世明. 玉米oxylipins信号途径关键基因的克隆及其在虫害诱导防御中的作用. 科学通报, 2005, 50: 2217-2225 |
| Xu T, Wang J B, Luo S M.Cloning of key genes involved in oxylipins signaling pathway and their roles in insect induced defense in maize.Chin Sci Bull, 2005, 50: 2217-2225 (in Chinese) | |
| [19] | Xu F, Zhao Y X, Wang F, Guo D L, Wei Y L, Huang X Z.Cloning of a vacuolar H+-pryophosphatase gene from emphemeral plant olimarabidopsispumila whose overexpression improve salt tolerance in tobacco.Afr J Biotechnol, 2013, 12: 6817-6825 |
| [20] | Horsch R, Fry J, Hoffmann N, Eichholtz D, Rogers S A, Fraley R.A simple and general method for transferring genes into plants.Science, 1985, 227: 1229-1231 |
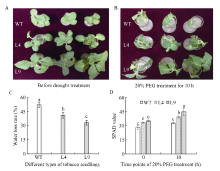
| [21] | Du H, Wang N, Cui F, Li X, Xiao J, Xiong L.Characterization of the beta-carotene hydroxylase gene DSM2 conferring drought and oxidative stress resistance by increasing xanthophylls and abscisic acid synthesis in rice. Plant Physiol, 2010, 154: 1304-1318 |
| [22] | 苍晶, 赵会杰. 植物生理学实验教程. 北京: 高等教育出版社, 2013. pp 151-153 |
| Cang J, Zhao H J.Experimental Guide of Plant Physiology. Beijing: Higher Education Press, 2013. pp 151-153 (in Chinese) | |
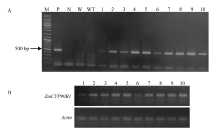
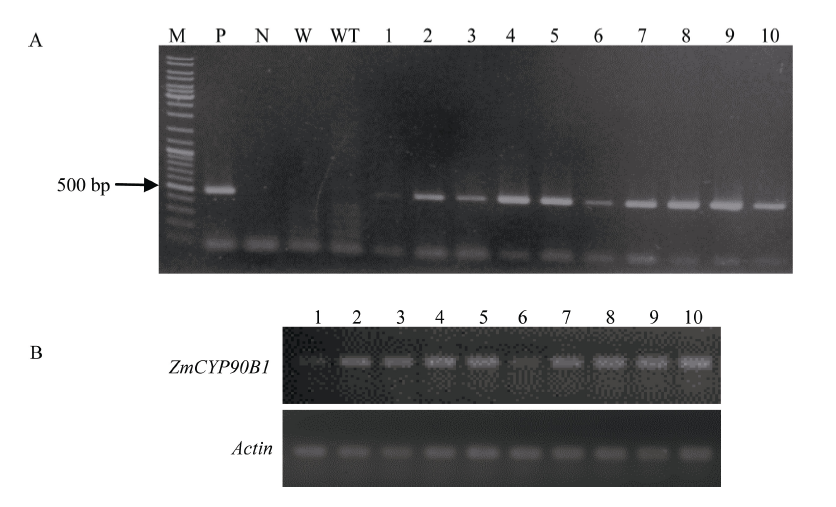
| [23] | Liu T, Zhang J, Wang M, Wang Z, Li G, Qu L, Wang G.Expression and functional analysis of ZmDWF4, an ortholog of Arabidopsis DWF4 from maize(Zea mays L.). Plant Cell Rep, 2007, 26: 2091-2099 |
| [24] | Divi U K, Krishna P.Overexpression of the brassinosteroid biosynthetic gene AtDWF4 in Arabidopsis seeds overcomes abscisic acid-induced inhibition of germination and increases cold tolerance in transgenic seedlings. Plant Growth Regul, 2010, 29: 385-393 |
| [25] | Wang Y, Chen Y F, Chang X H, Wang X, Li N, Gao Y F.Heterologous expression of Populus euphratica DWF4 (PeDWF4) enhanced tolerance to abiotic stress in tobacco plants. Genomics Appl Biol, 2017, Heterologous expression of Populus euphratica DWF4 (PeDWF4) enhanced tolerance to abiotic stress in tobacco plants. Genomics Appl Biol, 2017, (in Chinese with English abstract) |
| [26] | Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R.Reactive oxygen species homeostasis and signaling during drought and salinity stresses.Plant Cell Environ, 2010, 33: 453-457 |
| [27] | Kessler A, Baldwin I T.Plant response to insect herbivory: The emerging molecular analysis.Annu Rev Plant Biol, 2002, 53: 299-328 |
| [28] | Ryan C A.The systemin signaling pathway: differential activation of plant defensive genes.Biochim Biophy Acta, 2000, 1477: 112-121 |
| [29] | Li L, Li C, Lee G I, Howe G A.Distinct roles for jasmonate synthesis and action in the systemic wound response of tomato.Proc Natl Acad Sci USA, 2002, 99: 6416-6421 |
| [1] | WANG Dan-Dan, LIU Hong-Juan, WANG Hong-Xia, ZHANG Peng, SHI Chun-Yu. Cloning and functional analysis of the sweet potato sucrose transporter IbSUT3 [J]. Acta Agronomica Sinica, 2020, 46(7): 1120-1127. |
| [2] | ZHAI Yu-Shan, DENG Yu-Qing, DONG Meng, XU Qian, CHENG Guang-Yuan, PENG Lei, LIN Yan-Quan*,XU Jing-Sheng*. Cloning and Characterization of Light Harvesting Chlorophyll a/b-Binding Protein Coding Gene (ScLhca3) in Sugarcane [J]. Acta Agron Sin, 2016, 42(09): 1332-1341. |
| [3] | SU Wei-Hua**,LIU Feng**,HUANG Long,SU Ya-Chun,HUANG Ning,LING Hui,WU Qi-Bin,ZHANG Hua,QUE You-Xiong*. Cloning and Expression Analysis of a Ca2+/H+ Antiporter Gene from Sugarcane [J]. Acta Agron Sin, 2016, 42(07): 1074-1082. |
| [4] | LIU Feng**,SU Wei-Hua**,HUANG Long,XIAO Xin-Huan,HUANG Ning,LING Hui,SU Ya-Chun,ZHANG Hua,QUE You-Xiong. Isolation and Characterization of a Na+/H+Antiporter Gene from Sugarcane [J]. Acta Agron Sin, 2016, 42(04): 501-512. |
| [5] | HAO Jun-Jie,HU Yu-Wei,GUO Xiao-Qin,ZhAO Fu-An,JIA Xin-He,GUO Li-Juan,ZHANG Zhi-Xing,WANG Qing-Dong. Analysis of Resistance to Verticillium Wilt in Cotton by Reciprocal Grafting and Real-time Quantitative PCR [J]. Acta Agron Sin, 2013, 39(07): 1179-1186. |
| [6] | DI Ying, LEI Ting-Ting, YAN Fan, HUANG Kai-Meng, LI Xiao-Wei, ZHANG Qiang-Lin, ZHANG Hai-Jun, SU Lian-Tai, SUN Cuan, WANG Yang, LI Jing-Wen, WANG Qiang-Yu. Cloning and Expression of a Stress-induced GmPRP Gene in Soybean (Glycine max) [J]. Acta Agron Sin, 2011, 37(12): 2152-2157. |
| [7] | YANG Jing-Jing, LI Ya-Ning, LI Xing, LIU Da-Qun. Involvement of Heterotrimeric G Protein α and β Subunits in Defense Responses of Wheat to Puccinia triticina [J]. Acta Agron Sin, 2010, 36(12): 2028-2034. |
| [8] | MA Xiong-Feng,YU Chun-Ming,TANG Shou-Wei,ZHU Ai-Guo,WANG Yan-Zhou,ZHU Si-Yuan,. Cloning and Tissue Expression of Acting1 Gene in Different Fiber Development Phases of Ramie [Boehmeria nivea (Linn.) Gaud] [J]. Acta Agron Sin, 2010, 36(1): 101-108. |
| [9] | LIU Jian-Xin;YU Chun-Ming;TANG Shou-Wei;ZHU Ai-Guo;WANG Yan-Zhou;ZHU Si-Yuan;MA Xiong-Feng;XIONG He-Ping. Cloning and Tissue Expression of Important Enzyme Gene UGlcAE in Ramie Pectin Biosynthesis [J]. Acta Agron Sin, 2008, 34(11): 1938-1945. |
| [10] | YU Shun-Wu;LIU Hong-Yan;LUO Li-Jun. Analysis of Relative Gene Expression Using Different Real-time Quantitative PCR [J]. Acta Agron Sin, 2007, 33(07): 1214-1218. |
| [11] | CHEN Ying;SU Ning;XU Bo-Liang;GE Yi-Qiang;WANG Shu-Guang. Real-time Quantitative PCR Detection of Genetically Modified Maximizerâ Maize and YieldGardâ Maize [J]. Acta Agron Sin, 2004, 30(06): 602-607. |
|
||