欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (12): 2371-2378.doi: 10.3724/SP.J.1006.2021.01094
胡蕊洁( ), 杨向芸, 贾磊, 李玉如, 项月, 岳洁瑜, 王华忠*()
), 杨向芸, 贾磊, 李玉如, 项月, 岳洁瑜, 王华忠*()
HU Rui-Jie(), YANG Xiang-Yun, JIA Lei, LI Yu-Ru, XIANG Yue, YUE Jie-Yu, WANG Hua-Zhong*()
摘要:
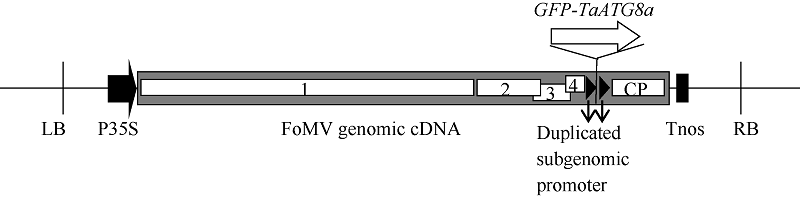
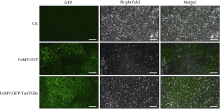
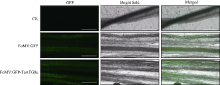
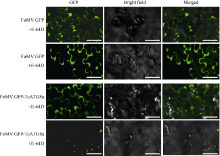
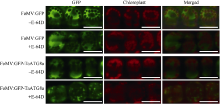
自噬相关因子ATG8定位于自噬结构的膜上, 荧光蛋白标记的ATG8在过表达细胞内所呈现的点状荧光常用于表征自噬结构和监测自噬活性。病毒介导的过表达(virus-mediated over-expression, VOX)是一种简便、快速制备目的基因过表达植株的技术。采用基于狗尾草花叶病毒FoMV的VOX技术(FoMV-VOX)在小麦植株中表达GFP标记的小麦ATG8家族成员TaATG8a, 建立小麦活体植株的自噬活性监测技术平台。构建了融合基因GFP-TaATG8a的FoMV-VOX载体, 采用Agroinfiltration方法在本氏烟草叶片中表达携带GFP-TaATG8a的FoMV基因组RNA和组装病毒粒子, 将烟草汁液中的病毒粒子摩擦接种于小麦幼苗植株叶片, 对接种植株叶片和根组织中的荧光信号进行观察和特征鉴定。结果表明, 采用FoMV-VOX技术在小麦植株上不仅可以实现GFP-TaATG8a在接种叶片中的高效表达, 还可以借助病毒的系统侵染实现该融合基因在未接种叶片和根组织中的高效表达。经饥饿处理激活自噬, 融合蛋白GFP-TaATG8a在植株叶表皮、叶肉以及根细胞中呈现表征自噬结构的点状荧光。采用FoMV-VOX技术获得的GFP-TaATG8a过表达植株可以应用于小麦多种组织类型中的自噬活性调节机制和生理功能研究。
| [1] |
Liu Y, Bassham D C. Autophagy: pathways for self-eating in plant cells. Annu Rev Plant Biol, 2012, 63:215-237.
doi: 10.1146/annurev-arplant-042811-105441 |
| [2] | Yang X, Bassham D C. New insight into the mechanism and function of autophagy in plant cells. Int Rev Cell Mol Biol, 2015, 320:1-40. |
| [3] |
Wang P, Mugume Y, Bassham D C. New advances in autophagy in plants: regulation, selectivity and function. Semin Cell Dev Biol, 2018, 80:113-122.
doi: 10.1016/j.semcdb.2017.07.018 |
| [4] |
Klionsky D J, Abdelmohsen K, Abe A, et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edn). Autophagy, 2016, 12:1-222.
doi: 10.1080/15548627.2015.1100356 pmid: 26799652 |
| [5] |
Bouton C, King R C, Chen H, Azhakanandam K, Bieri S, Hammond-Kosack K E, Kanyuka K. Foxtail mosaic virus: a viral vector for protein expression in cereals. Plant Physiol, 2018, 177:1352-1367.
doi: 10.1104/pp.17.01679 |
| [6] |
Dommes A B, Gross T, Herbert D B, Kivivirta K I, Becker A. Virus-induced gene silencing: empowering genetics in non-model organisms. J Exp Bot, 2019, 70:757-770.
doi: 10.1093/jxb/ery411 pmid: 30452695 |
| [7] |
Kant R, Dasgupta I. Gene silencing approaches through virus-based vectors: speeding up functional genomics in monocots. Plant Mol Biol, 2019, 100:3-18.
doi: 10.1007/s11103-019-00854-6 |
| [8] |
Lee W S, Hammond-Kosack K E, Kanyuka K. Barley stripe mosaic virus-mediated tools for investigating gene function in cereal plants and their pathogens: virus-induced gene silencing, host- mediated gene silencing, and virus-mediated over-expression of heterologous protein. Plant Physiol, 2012, 160:582-590.
doi: 10.1104/pp.112.203489 |
| [9] |
Liu N, Xie K, Jia Q, Zhao J, Chen T, Li H, Wei X, Diao X, Hong Y, Liu Y. Foxtail mosaic virus-induced gene silencing in monocot plants. Plant Physiol, 2016, 171:1801-1807.
doi: 10.1104/pp.16.00010 pmid: 27225900 |
| [10] | Mei Y, Beernink B M, Ellison E E, Konečná E, Neelakandan A K, Voytas D F, Whitham S A. Protein expression and gene editing in monocots using foxtail mosaic virus vectors. Plant Direct, 2019, 3:e00181. |
| [11] |
Li F, Vierstra R D. Autophagy: a multifaceted intracellular system for bulk and selective recycling. Trends Plant Sci, 2012, 17:526-537.
doi: 10.1016/j.tplants.2012.05.006 |
| [12] | Bassham D C. Function and regulation of macroautophagy in plants. Biochim Biophys Acta, 2009, 9:1397-1403. |
| [13] |
Nakatogawa H, Ichimura Y, Ohsumi Y. Atg8, a ubiquitin-like protein required for autophagosome formation, mediates membrane tethering and hemifusion. Cell, 2007, 130:165-178.
pmid: 17632063 |
| [14] | Izumi M, Hidema J, Wada S, Kondo E, Kurusu T, Kuchitsu K, Makino A, Ishida H. Establishment of monitoring methods for autophagy in rice reveals autophagic recycling of chloroplasts and root plastids during energy limitation. Plant Physiol, 2015, 167:1307-1320. |
| [15] |
Merkulova E A, Guiboileau A, Naya L, Masclaux-Daubresse C, Yoshimoto K. Assessment and optimization of autophagy monitoring methods in Arabidopsis roots indicate direct fusion of autophagosomes with vacuoles. Plant Cell Physiol, 2014, 55:715-726.
doi: 10.1093/pcp/pcu041 |
| [16] |
Pei D, Zhang W, Sun H, Wei X J, Yue J Y, Wang H Z. Identification of autophagy-related genes ATG4 and ATG8 from wheat ( Triticum aestivum L.) and profiling of their expression patterns responding to biotic and abiotic stresses. Plant Cell Rep, 2014, 33:1697-1710.
doi: 10.1007/s00299-014-1648-x |
| [17] |
Li K X, Liu Y N, Yu B J, Yang W W, Yue J Y, Wang H Z. Monitoring autophagy in wheat living cells by visualization of fluorescence protein-tagged ATG8. Plant Cell Tissue Organ Cult, 2018, 134:481-489.
doi: 10.1007/s11240-018-1437-2 |
| [1] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [2] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [3] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [4] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
| [5] | 刘运景, 郑飞娜, 张秀, 初金鹏, 于海涛, 代兴龙, 贺明荣. 宽幅播种对强筋小麦籽粒产量、品质和氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 716-725. |
| [6] | 马红勃, 刘东涛, 冯国华, 王静, 朱雪成, 张会云, 刘静, 刘立伟, 易媛. 黄淮麦区Fhb1基因的育种应用[J]. 作物学报, 2022, 48(3): 747-758. |
| [7] | 徐龙龙, 殷文, 胡发龙, 范虹, 樊志龙, 赵财, 于爱忠, 柴强. 水氮减量对地膜玉米免耕轮作小麦主要光合生理参数的影响[J]. 作物学报, 2022, 48(2): 437-447. |
| [8] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [9] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
| [10] | 马博闻, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑, 姜东. 花前渍水锻炼调控花后小麦耐渍性的生理机制研究[J]. 作物学报, 2022, 48(1): 151-164. |
| [11] | 孟颖, 邢蕾蕾, 曹晓红, 郭光艳, 柴建芳, 秘彩莉. 小麦Ta4CL1基因的克隆及其在促进转基因拟南芥生长和木质素沉积中的功能[J]. 作物学报, 2022, 48(1): 63-75. |
| [12] | 韦一昊, 于美琴, 张晓娇, 王露露, 张志勇, 马新明, 李会强, 王小纯. 小麦谷氨酰胺合成酶基因可变剪接分析[J]. 作物学报, 2022, 48(1): 40-47. |
| [13] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [14] | 罗江陶, 郑建敏, 蒲宗君, 范超兰, 刘登才, 郝明. 四倍体小麦与六倍体小麦杂种的染色体遗传特性[J]. 作物学报, 2021, 47(8): 1427-1436. |
| [15] | 王艳朋, 凌磊, 张文睿, 王丹, 郭长虹. 小麦B-box基因家族全基因组鉴定与表达分析[J]. 作物学报, 2021, 47(8): 1437-1449. |
|
||