欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (8): 1146-1156.doi: 10.3724/SP.J.1006.2020.94198
冯韬1,2,谭晖1,2,官梅2,官春云2,*( )
)
FENG Tao1,2,TAN Hui1,2,GUAN Mei2,GUAN Chun-Yun2,*()
摘要:
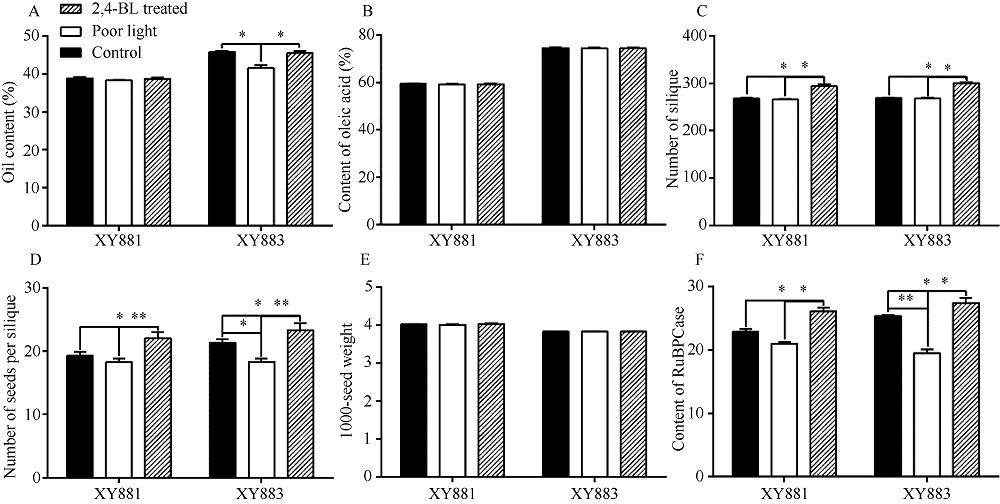
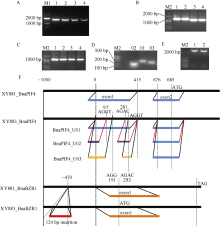
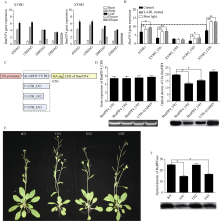
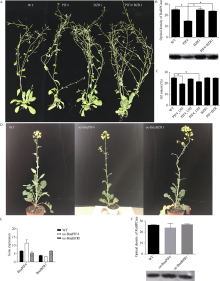
甘蓝型油菜品系XY881和XY883是湘油15辐照诱变后连续自交筛选的2个种子含油量、光合效率和弱光敏感性等有明显差别的子代品系。分别从XY881和XY883中克隆了芸薹素唑抗性因子1 (brassinazole-resistant 1, BnaBZR1) 和光敏色素互作因子4 (phytochrome interacting factor 4, BnaPIF4)基因并进行了序列结构、表达和功能分析。结果表明, XY883的BnaBZR1和BnaPIF4基因存在结构变异, 引起表达和调控模式的差异。XY883中BnaBZR1的启动子具有124 bp的富含A/T的插入序列, 且XY883具有比XY881高的BnaBZR1表达, 并且在弱光和2,4-表油菜素内酯(2,4-BL)诱导下具有较少的表达变化。XY883中BnaPIF4的5'-UTR区域存在可变剪接, 形成长度分别为424 bp (U01)、239 bp (U02)和332 bp (U03)的3种5'-UTR, 在弱光和2,4-BL诱导下, XY883中3种可变剪接的BnaPIF4转录产物的变化不一致。将BnaPIF4的3个5'-UTR与CDS分别组合转化拟南芥后其表达在转录水平无明显差异, 但蛋白翻译存在明显差异, 表明BnaPIF4的5'-UTR变异影响其翻译过程。转BnaPIF4基因拟南芥出现株高增加、叶片狭长且光合作用下降的表型, 共转化BnaBZR1能减弱BnaPIF4造成的光合作用下降; 转BnaPIF4和BnaBZR1基因对油菜的影响与拟南芥相似, 但表型不如拟南芥明显, 表明BnaPIF4是油菜光合作用的负调控因子, 而BnaBZR1可对BnaPIF4的光合负调控产生拮抗; 这与XY881和XY883中两基因表达调控模式及其光合表型相吻合。
| [1] |
Oh E, Zhu J Y, Wang Z Y. Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nat Cell Biol, 2012,14:802-809.
doi: 10.1038/ncb2545 pmid: 22820378 |
| [2] |
Bai M Y, Shang J X, Oh E, Fan M, Bai Y, Zentella R, Sun T P, Wang Z Y. Brassinosteroid, gibberellin and phytochrome impinge on a common transcription module in Arabidopsis. Nat Cell Biol, 2012,14:810-817.
doi: 10.1038/ncb2546 pmid: 22820377 |
| [3] |
Gudesblat G E, Russinova E. Plants grow on brassinosteroids. Curr Opin Plant Biol, 2011,14:530-537.
pmid: 21802346 |
| [4] | Ryu H, Hwang I. Brassinosteroids in plant developmental signaling networks. J Plant Biol, 2013,56:267-273. |
| [5] | Wang W, Bai M Y, Wang Z Y. The brassinosteroid signaling network—a paradigm of signal integration. Curr Opin Plant Biol, 2014,21:147-153. |
| [6] | USDA FAS (Foreign Agricultural Service), 2016. Oilseeds: World Markets and Trade. https://www.fas.usda.gov/data/oilseeds-world-markets-and-trade. |
| [7] |
Chalhoub B, Denoeud F, Liu S, Parkin I A, Tang H, Wang X, Corréa M. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science, 2014,345:950-953.
doi: 10.1126/science.1253435 pmid: 25146293 |
| [8] |
Mølmann J A, Hagen S F, Bengtsson G B, Johansen T J. Influence of high latitude light conditions on sensory quality and contents of health and sensory-related compounds in swede roots (Brassica napus L. ssp. rapifera Metzg.). J Sci Food Agric, 2018,98:1117-1123.
doi: 10.1002/jsfa.8562 pmid: 28732144 |
| [9] | 冯韬, 官春云. 甘蓝型油菜光敏色素互作因子4 (BnaPIF4)基因克隆和功能分析. 作物学报, 2019,45:204-213. |
| Feng T, Guan C Y. Cloning and characterization of phytochrome interacting factor 4 (BnaPIF4) gene from Brassica napus L. Acta Agron Sin, 2019,45:204-213 (in Chinese with English abstract). | |
| [10] |
Wei F, Gao G Z, Wang X F, Dong X Y, Li P P, Hua W, Wang X, Wu X M, Chen H. Quantitative determination of oil content in small quantity of oilseed rape by ultrasound-assisted extraction combined with gas chromatography. Ultrason Sonochem, 2008,15:938-942.
pmid: 18504157 |
| [11] | 冯韬, 谭晖, 徐江林, 官春云. 油菜素内酯在不同生育期对两品系甘蓝型油菜的生长调控. 中国油料作物学报, 2019,41:904-913. |
| Feng T, Tan H, Xu J L, Guan C Y. Epibrassinolide regulation on oilseed rape (Brassica napus L.) in different period. Chin J Oil Crop Sci, 2019,41:904-913 (in Chinese with English abstract). | |
| [12] |
Wang Z Y, Nakano T, Gendron J, He J X, Chen M, Vafeados D, Yang Y L, Fujioka S, Yoshida S, Asami T, Chory J. Nuclear-localized BZR1 m llklediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Dev Cell, 2002,2:505-513.
doi: 10.1016/s1534-5807(02)00153-3 pmid: 11970900 |
| [13] |
Casson S A, Franklin K A, Gray J E, Grierson C S, Whitelam G C, Hetherington A M. Phytochrome B and PIF4 regulate stomatal development in response to light quantity. Curr Biol, 2009,19:229-234.
doi: 10.1016/j.cub.2008.12.046 pmid: 19185498 |
| [14] |
Kumar S V, Lucyshyn D, Jaeger K E, Alós E, Alvey E, Harberd N P, Wigge P A. Transcription factor PIF4 controls the thermosensory activation of flowering. Nature, 2012,484:242-245.
doi: 10.1038/nature10928 pmid: 22437497 |
| [15] |
Lucas M D, Davière J M, Mariana R F, Pontin M, Manuel J I P, Lorrain S, Fankhauser C, Blázquez M A, Titarenko E, Prat S. A molecular framework for light and gibberellin control of cell elongation. Nature, 2008,451:480-484.
doi: 10.1038/nature06520 pmid: 18216857 |
| [16] |
Stella B G, Miguel D L, Cristina M, Ana E R, Davière J M, Prat S. BR-dependent phosphorylation modulates PIF4 transcriptional activity and shapes diurnal hypocotyl growth. Genes Dev, 2014,28:1681-1694.
doi: 10.1101/gad.243675.114 pmid: 25085420 |
| [17] |
Franklin K A, Lee S H, Patel D, Kumar S V, Spartz A K, Gu C, Ye S Q, Yu P, Breen G, Cohen J D, Wigge P A, Gray W M. PHYTOCHROME-NTERACTING FACTOR 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc Natl Acad Sci USA, 2011,108:20231-20235.
doi: 10.1073/pnas.1110682108 pmid: 22123947 |
| [18] | 冯韬, 官春云. 甘蓝型油菜芸薹素唑耐受因子(BnaBZR1/ BnaBES1)全长CDS克隆与生物信息学分析. 作物学报, 2018,44:1793-1801. |
| Feng T, Guan C Y. Cloning and characterization of brassinazole-resistant (BnaBZR1 and BnaBES1) CDS from Brassica napus L. Acta Agron Sin, 2018,44:1793-1801 (in Chinese with English abstract). |
| [1] | 陈松余, 丁一娟, 孙峻溟, 黄登文, 杨楠, 代雨涵, 万华方, 钱伟. 甘蓝型油菜BnCNGC基因家族鉴定及其在核盘菌侵染和PEG处理下的表达特性分析[J]. 作物学报, 2022, 48(6): 1357-1371. |
| [2] | 秦璐, 韩配配, 常海滨, 顾炽明, 黄威, 李银水, 廖祥生, 谢立华, 廖星. 甘蓝型油菜耐低氮种质筛选及绿肥应用潜力评价[J]. 作物学报, 2022, 48(6): 1488-1501. |
| [3] | 袁大双, 邓琬玉, 王珍, 彭茜, 张晓莉, 姚梦楠, 缪文杰, 朱冬鸣, 李加纳, 梁颖. 甘蓝型油菜BnMAPK2基因的克隆及功能分析[J]. 作物学报, 2022, 48(4): 840-850. |
| [4] | 黄成, 梁晓梅, 戴成, 文静, 易斌, 涂金星, 沈金雄, 傅廷栋, 马朝芝. 甘蓝型油菜BnAPs基因家族成员全基因组鉴定及分析[J]. 作物学报, 2022, 48(3): 597-607. |
| [5] | 王瑞, 陈雪, 郭青青, 周蓉, 陈蕾, 李加纳. 甘蓝型油菜白花基因InDel连锁标记开发[J]. 作物学报, 2022, 48(3): 759-769. |
| [6] | 王艳花, 刘景森, 李加纳. 整合GWAS和WGCNA筛选鉴定甘蓝型油菜生物产量候选基因[J]. 作物学报, 2021, 47(8): 1491-1510. |
| [7] | 李杰华, 端群, 史明涛, 吴潞梅, 柳寒, 林拥军, 吴高兵, 范楚川, 周永明. 新型抗广谱性除草剂草甘膦转基因油菜的创制及其鉴定[J]. 作物学报, 2021, 47(5): 789-798. |
| [8] | 唐鑫, 李圆圆, 陆俊杏, 张涛. 甘蓝型油菜温敏细胞核雄性不育系160S花药败育的形态学特征和细胞学研究[J]. 作物学报, 2021, 47(5): 983-990. |
| [9] | 周新桐, 郭青青, 陈雪, 李加纳, 王瑞. GBS高密度遗传连锁图谱定位甘蓝型油菜粉色花性状[J]. 作物学报, 2021, 47(4): 587-598. |
| [10] | 李书宇, 黄杨, 熊洁, 丁戈, 陈伦林, 宋来强. 甘蓝型油菜早熟性状QTL定位及候选基因筛选[J]. 作物学报, 2021, 47(4): 626-637. |
| [11] | 张春, 赵小珍, 庞承珂, 彭门路, 王晓东, 陈锋, 张维, 陈松, 彭琦, 易斌, 孙程明, 张洁夫, 傅廷栋. 甘蓝型油菜千粒重全基因组关联分析[J]. 作物学报, 2021, 47(4): 650-659. |
| [12] | 唐婧泉, 王南, 高界, 刘婷婷, 文静, 易斌, 涂金星, 傅廷栋, 沈金雄. 甘蓝型油菜SnRK基因家族生物信息学分析及其与种子含油量的关系[J]. 作物学报, 2021, 47(3): 416-426. |
| [13] | 蒙姜宇, 梁光伟, 贺亚军, 钱伟. 甘蓝型油菜耐盐和耐旱相关性状的QTL分析[J]. 作物学报, 2021, 47(3): 462-471. |
| [14] | 魏丽娟, 申树林, 黄小虎, 马国强, 王曦彤, 杨怡玲, 李洹东, 王书贤, 朱美晨, 唐章林, 卢坤, 李加纳, 曲存民. 锌胁迫下甘蓝型油菜发芽期下胚轴长的全基因组关联分析[J]. 作物学报, 2021, 47(2): 262-274. |
| [15] | 李倩, Nadil Shah, 周元委, 侯照科, 龚建芳, 刘珏, 尚政伟, 张磊, 战宗祥, 常海滨, 傅廷栋, 朴钟云, 张椿雨. 抗根肿病甘蓝型油菜新品种华油杂62R的选育[J]. 作物学报, 2021, 47(2): 210-223. |
|
||