欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (12): 2394-2406.doi: 10.3724/SP.J.1006.2021.04259
解盼1,4( ), 刘蔚1, 康郁1, 华玮1, 钱论文1,2,3, 官春云1,2,3,*, 何昕1,2,3,*
), 刘蔚1, 康郁1, 华玮1, 钱论文1,2,3, 官春云1,2,3,*, 何昕1,2,3,*
XIE Pan1,4(), LIU Wei1, KANG Yu1, HUA Wei1, QIAN Lun-Wen1,2,3, GUAN Chun-Yun1,2,3,*, HE Xin1,2,3,*
摘要:
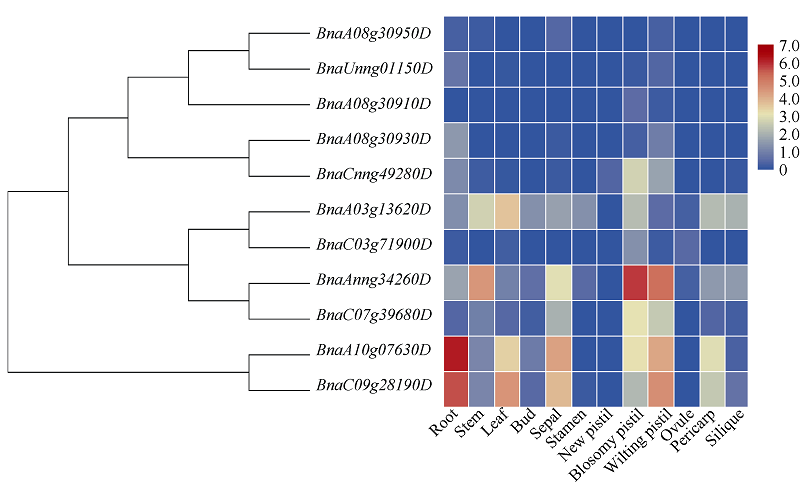
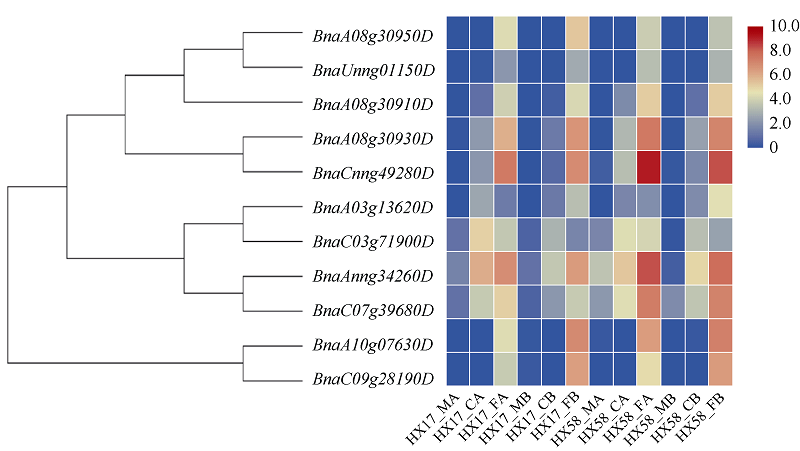
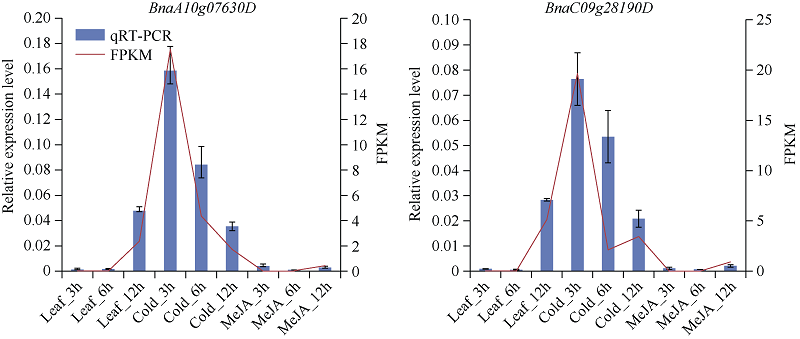
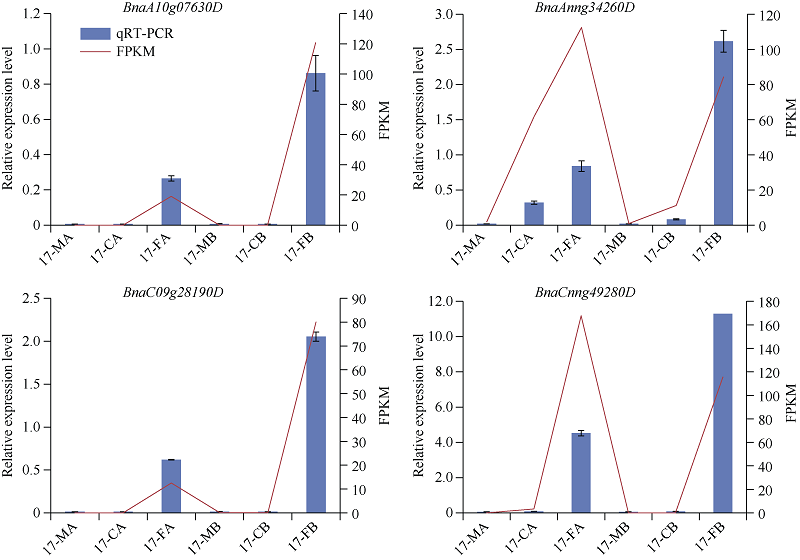
低温是影响植物生长发育的重要环境胁迫因子, ICE1 (inducer of CBF expression1)-CBF (C-repeat (CRT)-binding factors)-COR (cold responsive)在植物响应低温胁迫的信号途径中具有重要作用。为探究CBF (C-repeat-binding factors)基因在甘蓝型油菜(Brassica napus L.)中的进化以及在低温胁迫应答反应中的功能, 本研究以4个拟南芥CBF基因为基础序列, 鉴定出11个甘蓝型油菜、6个白菜和5个甘蓝CBF基因, 并对它们的分子特性、蛋白保守结构域和系统进化树、基因结构及基因染色体分布、甘蓝型油菜CBF基因组织表达模式以及不同逆境和激素处理下的表达模式进行系统的比较分析。结果表明, 在11个甘蓝型油菜CBF基因中, 可分为亚组I (CBF1/2/3)和亚组II (CBF4) 2个亚组。转录组测序结果表明, 所有甘蓝型油菜CBF基因受低温诱导表达, 其中亚组Ib的4个基因对冷胁迫响应迅速且持续时间长, 亚组II中2个基因对冷胁迫响应表达较弱, 但它们在根中表达量明显高于叶片, 并且参与盐胁迫和渗透胁迫响应; 甘蓝型油菜中CBF基因家族对冻害响应更强烈, 其中亚组I中的BnaA08g30930D、BnaCnng49280D、BnaAnng34260D、BnaC07g39680D和亚组II中BnaA10g07630D、BnaC09g28190D对冻害响应尤为显著。本研究将为进一步了解甘蓝型油菜CBF家族基因的生物学功能及其对低温尤其是冻害响应奠定基础, 同时为CBF家族基因在其他物种中的生物信息学研究提供参考。
| [1] | 陈明, 叶川, 吕伟生, 肖国滨, 应国勇, 王瑞平, 刘桅, 郑伟. 稻稻油三熟制下油菜早熟高产品种的筛选. 江苏农业科学, 2018, 46(17):61-64. |
| Chen M, Ye C, Lyu W S, Xiao G B, Ying G Y, Wang D P, Liu Z, Zheng W. Screening of early-maturing and high-yielding rapeseed varieties under the three-cropping system of rice-rice-oil. Jiangsu Agric Sci, 2018, 46(17):61-64 (in Chinese with English abstract). | |
| [2] | 张尧锋, 余华胜, 曾孝元, 林宝刚, 华水金, 张冬青, 傅鹰. 早熟甘蓝型油菜研究进展及其应用. 植物遗传资源学报, 2019, 20:258-266. |
| Zhang Y F, Yu H S, Zeng X Y, Lin B G, Hua S J, Zhang D Q, Fu Y. Progress and application of early maturity in rapeseed ( Brassica napus L.). J Plant Genet Resour, 2019, 20:258-266 (in Chinese with English abstract). | |
| [3] | 王必庆, 王国槐. 油菜早熟性研究进展. 作物研究, 2009, 23(5):336-338. |
| Wang B Q, Wang G H. Research progress on early maturity of rape. Crop Res, 2009, 23:336-338 (in Chinese with English abstract). | |
| [4] | 张学昆, 张春雷, 廖星, 王汉中. 2008年长江流域油菜低温冻害调查分析. 中国油料作物学报, 2008, 30:122-126. |
| Zhang X K, Zhang C L, Liao X, Wang H Z. Investigation on 2008' low temperature and freeze injure on winter rape along Yangtze River. Chin J Oil Crop Sci, 2008, 30:122-126 (in Chinese with English abstract). | |
| [5] | 涂玉琴, 戴兴临. 花期低温阴雨对甘蓝型油菜产量和种子含油量的影响. 中国油料作物学报, 2011, 33:470-475. |
| Tu Y Q, Dai X L. Effects of continuous low temperature overcast and rainy weather on yield and oil content of Brassica napus during flowering stage. Chin J Oil Crop Sci, 2011, 33:470-475 (in Chinese with English abstract). | |
| [6] |
Meshi T, Iwabuchi M. Plant transcription factors. Plant Cell Physiol, 1995, 36:1405-1420.
pmid: 8589926 |
| [7] |
Stockinger E J, Gilmour S J, Thomashow M F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc Natl Acad Sci USA, 1997, 94:1035-1040.
doi: 10.1073/pnas.94.3.1035 |
| [8] |
Gilmour S J, Zarka D G, Stockinger E J, Salazar M P, Houghton J M, Thomashow M F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J, 1998, 16:433-442.
pmid: 9881163 |
| [9] |
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell, 1998, 10:1391-1406.
pmid: 9707537 |
| [10] | Ding Y L, Shi Y T, Yang S H. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol, 2019, 20:1690-1704. |
| [11] |
Haake V, Cook D, Riechmann J L, Pineda O, Thomashow M F, Zhang J Z. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol, 2002, 130:639-648.
doi: 10.1104/pp.006478 |
| [12] |
Jaglo-Ottosen K R, Gilmour S J, Zarka D G, Schabenberger O, Thomashow M F, Jaglo-Ottosen K R, Gilmour S J, Zarka D G. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science, 1998. 280:104-106.
pmid: 9525853 |
| [13] |
Novillo F, Medina J, Salinas J. Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon. Proc Natl Acad Sci USA, 2007, 104:21002-21007.
doi: 10.1073/pnas.0705639105 |
| [14] |
Park S, Lee C M, Doherty C J, Gilmour S J, Kim Y, Thomashow M F. Regulation of the Arabidopsis CBF regulon by a complex low‐temperature regulatory network. Plant J, 2015, 82:193-207.
doi: 10.1111/tpj.2015.82.issue-2 |
| [15] |
Jia Y X, Ding Y L, Shi Y T, Zhang X Y, Gong Z Z, Yang S H. The cbfs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis. New Phytol, 2016, 212:345-353.
doi: 10.1111/nph.2016.212.issue-2 |
| [16] |
Zhao C Z, Zhang Z J, Xie S J, Si T, Li Y Y, Zhu J K. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis. Plant Physiol, 2016, 171:2744-2759.
doi: 10.1104/pp.16.00533 |
| [17] |
Yan L, Shah T, Cheng Y, Lyu Y, Zhang X K, Zou X L. Physiological and molecular responses to cold stress in rapeseed ( Brassica napus L.). J Integr Agric, 2019, 18:2742-2752.
doi: 10.1016/S2095-3119(18)62147-1 |
| [18] |
Karimi M, Ebadi A, Mousavi S A, Salami S A, Zarei A. Comparison of CBF1, CBF2, CBF3 and CBF4 expression in some grapevine cultivars and species under cold stress. Sci Hortic, 2015, 197:521-526.
doi: 10.1016/j.scienta.2015.10.011 |
| [19] | An D, Ma Q X, Yan W, Zhou W Z, Liu G H, Zhang P. Divergent regulation of CBF regulon on cold tolerance and plant phenotype in cassava overexpressing Arabidopsis CBF3 gene. Front Plant Sci, 2016, 7:1866. |
| [20] | 甄伟, 陈溪, 孙思洋, 胡鸢雷, 林忠平. 冷诱导基因的转录因子CBF1转化油菜和烟草及抗寒鉴定. 自然科学进展, 2002, 10:1104-1109. |
| Zhen W, Chen X, Sun S Y, Hu Y L, Lin Z P. Transformation of cold-induced gene transcription factor CBF1 into rape and tobacco and identification of cold tolerance. Prog Nat Sci, 2002, 10:1104-1109 (in Chinese with English abstract). | |
| [21] |
Hsieh T, Lee J T, Charng Y Y, Chan M T. Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced resistance to water deficit stress. Plant Physiol, 2002, 130:618-626.
doi: 10.1104/pp.006783 |
| [22] | 金建凤, 高强, 陈勇, 王君晖. 转移拟南芥CBF1基因引起水稻植株脯氨酸含量提高. 细胞生物学杂志, 2005, 27(1):73-76. |
| Jin J F, Gao Q, Chen Y, Wang J H. Transfer of Arabidopsis CBFl gene leads to increased proline contents in rice plants. Chin J Cell Biol, 2005, 27(1):73-76 (in Chinese with English abstract). | |
| [23] |
Ito Y, Katsura K, Maruyama K, Taji T, Kobayashi M, Seki M. Shinozaki K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol, 2006, 47:141-153.
doi: 10.1093/pcp/pci230 |
| [24] |
He X, Xie S, Xie P, Yao M, Liu W, Qin L W, Liu Z S, Zheng M, Liu H F, Guan M, Hua W. Genome-wide identification of stress-associated proteins ( SAP) with A20/AN1 zinc finger domains associated with abiotic stresses responses in Brassica napus. Environ Exp Bot, 2019, 165:108-119.
doi: 10.1016/j.envexpbot.2019.05.007 |
| [25] | He X, Ni X C, Xie P, Liu W, Yao M, Kang Y, Qin L W, Hua W. Comparative transcriptome analyses revealed conserved and novel responses to cold and freezing stress in Brassica napus L. G3: Genes Genom Genet, 2019, 9:2723-2737. |
| [26] |
Sudhir K, Glen S, Koichiro T. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol, 2016, 33:1870-1874.
doi: 10.1093/molbev/msw054 pmid: 27004904 |
| [27] |
Smith T F, Waterman M S. Identification of common molecular subsequences. J Mol Biol, 1981, 147:195-197.
pmid: 7265238 |
| [28] |
Savitch L V, Allard G, Seki M, Robert L S, Tinker N A, Huner N P, Shinozaki K, Singh J. The effect of overexpression of two Brassica CBF/DREB1-like transcription factors on photosynthetic capacity and freezing tolerance in Brassica napus. Plant Cell Physiol, 2005, 46:1525-1539.
pmid: 16024910 |
| [29] |
Shi Y, Ding Y, Yang S. Molecular regulation of CBF signaling in cold acclimation. Trends Plant Sci, 2018, 23:623-637.
doi: 10.1016/j.tplants.2018.04.002 |
| [30] |
Jaglo K R, Kleff S, Amundsen K L, Zhang X, Haake V, Zhang J Z, Deits T, Thomashow M F. Components of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol, 2001, 127:910-917.
pmid: 11706173 |
| [31] |
Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki K. A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought and low-temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol, 2004, 45:346-350.
pmid: 15047884 |
| [32] |
Qin F, Sakuma Y, Li J, Liu Q, Li Y Q, Shinozaki K, Yamaguchi-Shinozaki K. Cloning and functional analysis of a novel DREB1/CBF transcription factor involved in cold-responsive gene expression in Zea mays L. Plant Cell Physiol, 2004, 45:1042-1052.
doi: 10.1093/pcp/pch118 |
| [33] | Canella D, Gilmour S J, Kuhn L A, Thomashow M F. DNA binding by the Arabidopsis CBF1 transcription factor requires the PKKP/RAGRxKFxETRHP signature sequence. Biochim Biophys Acta, 2010, 1799:454-462. |
| [34] |
Medina J, Bargues M, Terol J, Perez-Alonso M, Salinas J. The Arabidopsis CBF gene family is composed of three genes encoding AP2 domain-containing proteins whose expression Is regulated by low temperature but not by abscisic acid or dehydration. Plant Physiol, 1999, 119:463-470.
pmid: 9952441 |
| [35] |
Gilmour S J, Sebolt A M, Salazar M P, Everard J D, Thomashow M F. Overexpression of the Arabidopsis CBF3 transcriptional activator mimics multiple biochemical changes associated with cold acclimation. Plant Physiol, 2000, 124:1854-1865.
pmid: 11115899 |
| [36] |
Zhang X, Fowler S, Cheng H M, Lou Y, Rhee S Y, Stockinger E J, Thomashow M F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing-tolerant Arabidopsis. Plant J, 2004, 39:905-919.
pmid: 15341633 |
| [1] | 陈松余, 丁一娟, 孙峻溟, 黄登文, 杨楠, 代雨涵, 万华方, 钱伟. 甘蓝型油菜BnCNGC基因家族鉴定及其在核盘菌侵染和PEG处理下的表达特性分析[J]. 作物学报, 2022, 48(6): 1357-1371. |
| [2] | 秦璐, 韩配配, 常海滨, 顾炽明, 黄威, 李银水, 廖祥生, 谢立华, 廖星. 甘蓝型油菜耐低氮种质筛选及绿肥应用潜力评价[J]. 作物学报, 2022, 48(6): 1488-1501. |
| [3] | 袁大双, 邓琬玉, 王珍, 彭茜, 张晓莉, 姚梦楠, 缪文杰, 朱冬鸣, 李加纳, 梁颖. 甘蓝型油菜BnMAPK2基因的克隆及功能分析[J]. 作物学报, 2022, 48(4): 840-850. |
| [4] | 冯亚, 朱熙, 罗红玉, 李世贵, 张宁, 司怀军. 马铃薯StMAPK4响应低温胁迫的功能解析[J]. 作物学报, 2022, 48(4): 896-907. |
| [5] | 黄成, 梁晓梅, 戴成, 文静, 易斌, 涂金星, 沈金雄, 傅廷栋, 马朝芝. 甘蓝型油菜BnAPs基因家族成员全基因组鉴定及分析[J]. 作物学报, 2022, 48(3): 597-607. |
| [6] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [7] | 王瑞, 陈雪, 郭青青, 周蓉, 陈蕾, 李加纳. 甘蓝型油菜白花基因InDel连锁标记开发[J]. 作物学报, 2022, 48(3): 759-769. |
| [8] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [9] | 薛晓梦, 吴洁, 王欣, 白冬梅, 胡美玲, 晏立英, 陈玉宁, 康彦平, 王志慧, 淮东欣, 雷永, 廖伯寿. 低温胁迫对普通和高油酸花生种子萌发的影响[J]. 作物学报, 2021, 47(9): 1768-1778. |
| [10] | 王艳花, 刘景森, 李加纳. 整合GWAS和WGCNA筛选鉴定甘蓝型油菜生物产量候选基因[J]. 作物学报, 2021, 47(8): 1491-1510. |
| [11] | 李杰华, 端群, 史明涛, 吴潞梅, 柳寒, 林拥军, 吴高兵, 范楚川, 周永明. 新型抗广谱性除草剂草甘膦转基因油菜的创制及其鉴定[J]. 作物学报, 2021, 47(5): 789-798. |
| [12] | 唐鑫, 李圆圆, 陆俊杏, 张涛. 甘蓝型油菜温敏细胞核雄性不育系160S花药败育的形态学特征和细胞学研究[J]. 作物学报, 2021, 47(5): 983-990. |
| [13] | 周新桐, 郭青青, 陈雪, 李加纳, 王瑞. GBS高密度遗传连锁图谱定位甘蓝型油菜粉色花性状[J]. 作物学报, 2021, 47(4): 587-598. |
| [14] | 李书宇, 黄杨, 熊洁, 丁戈, 陈伦林, 宋来强. 甘蓝型油菜早熟性状QTL定位及候选基因筛选[J]. 作物学报, 2021, 47(4): 626-637. |
| [15] | 张春, 赵小珍, 庞承珂, 彭门路, 王晓东, 陈锋, 张维, 陈松, 彭琦, 易斌, 孙程明, 张洁夫, 傅廷栋. 甘蓝型油菜千粒重全基因组关联分析[J]. 作物学报, 2021, 47(4): 650-659. |
|
||