欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (12): 3227-3238.doi: 10.3724/SP.J.1006.2023.24285
牛志远1,2, 秦超1, 刘军1, 王海泽2,*( ), 李宏宇1,*()
), 李宏宇1,*()
NIU Zhi-Yuan1,2, QIN Chao1, LIU Jun1, WANG Hai-Ze2,*(), LI Hong-Yu1,*()
摘要:
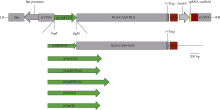
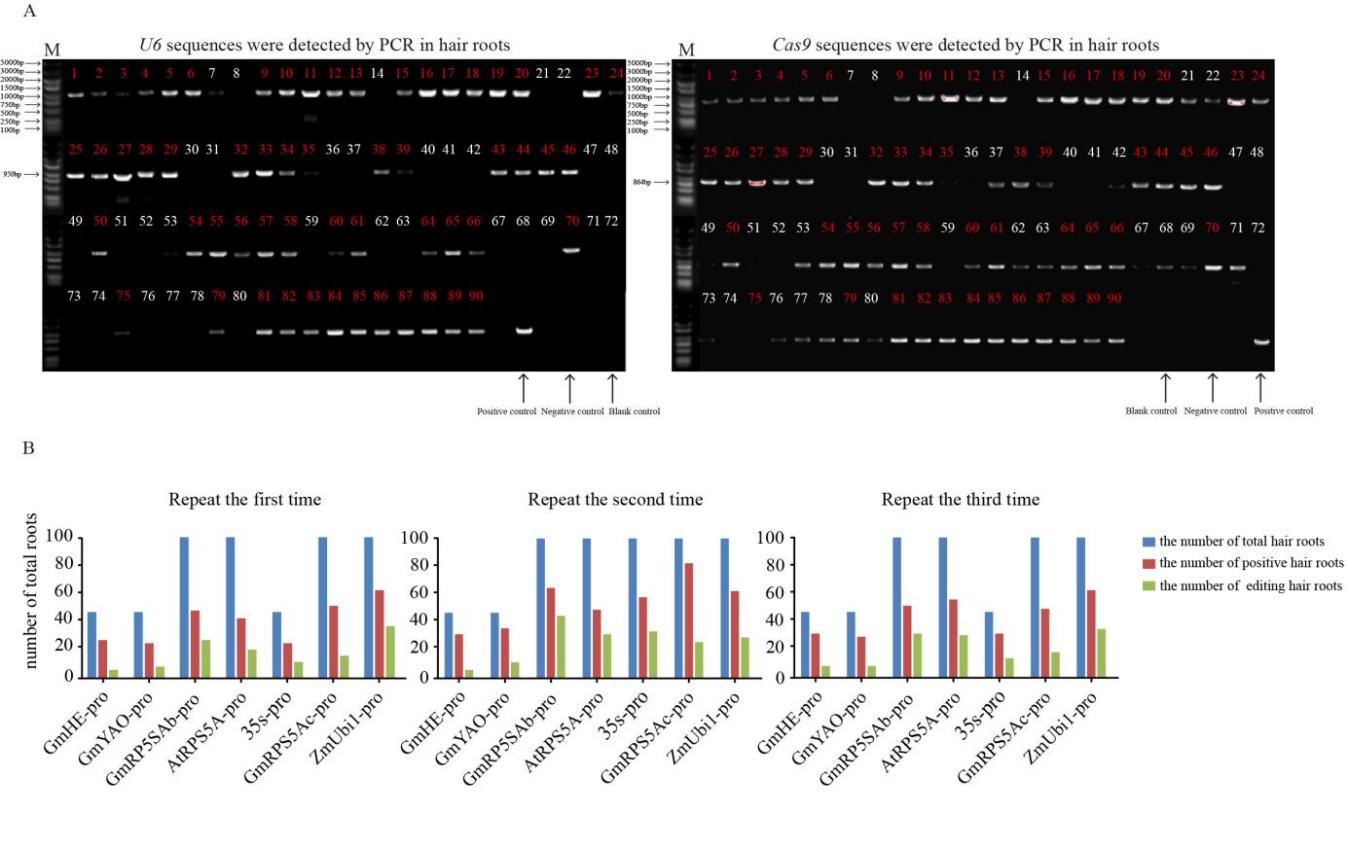
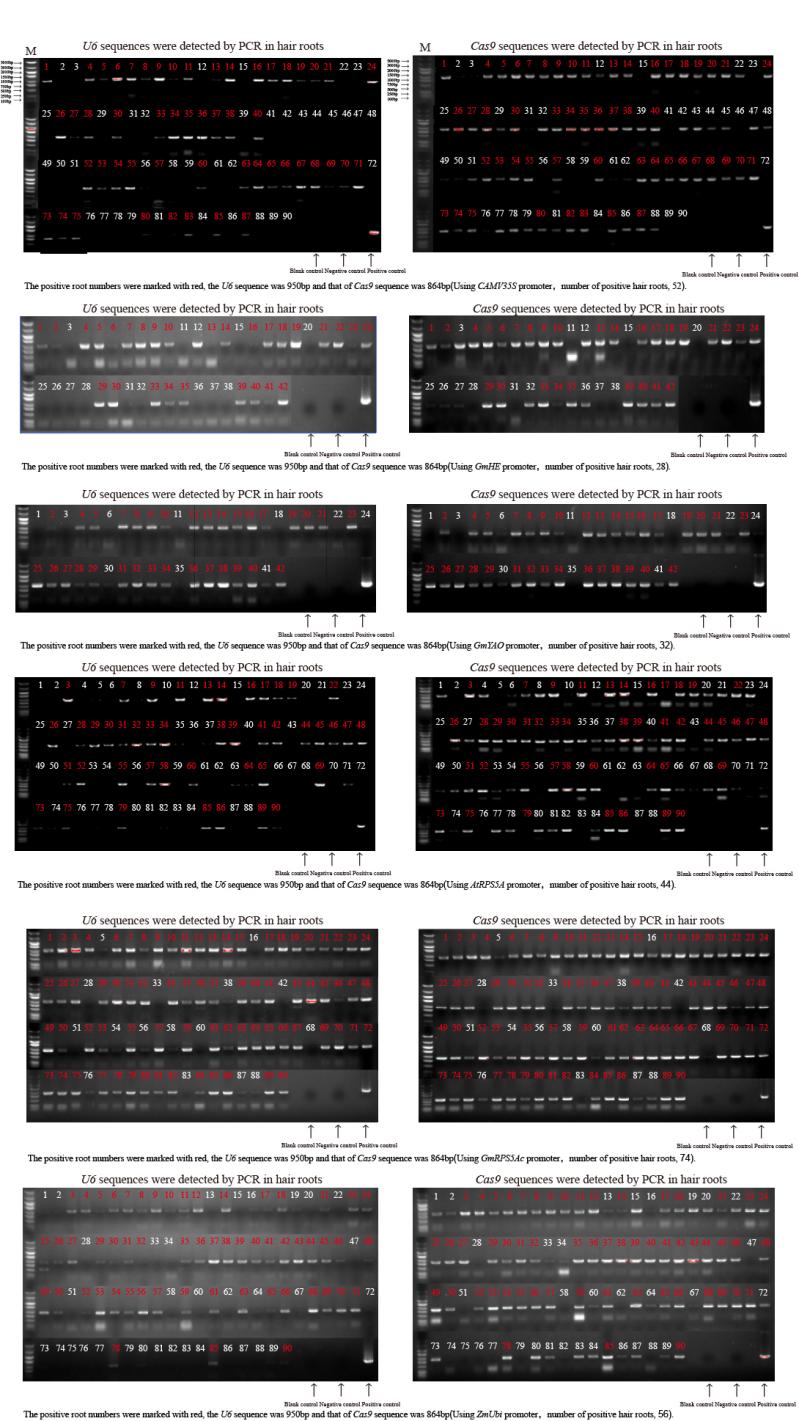
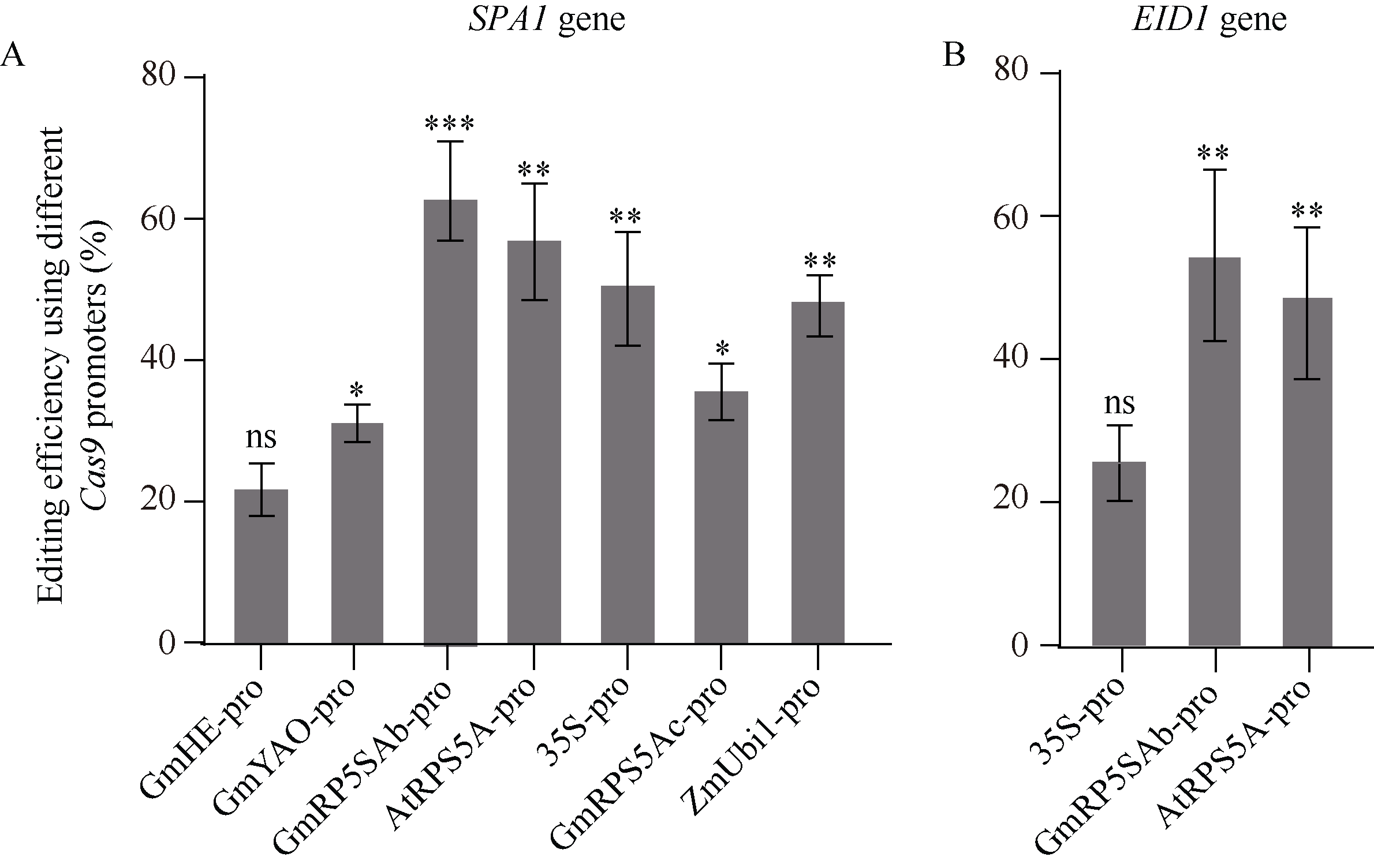
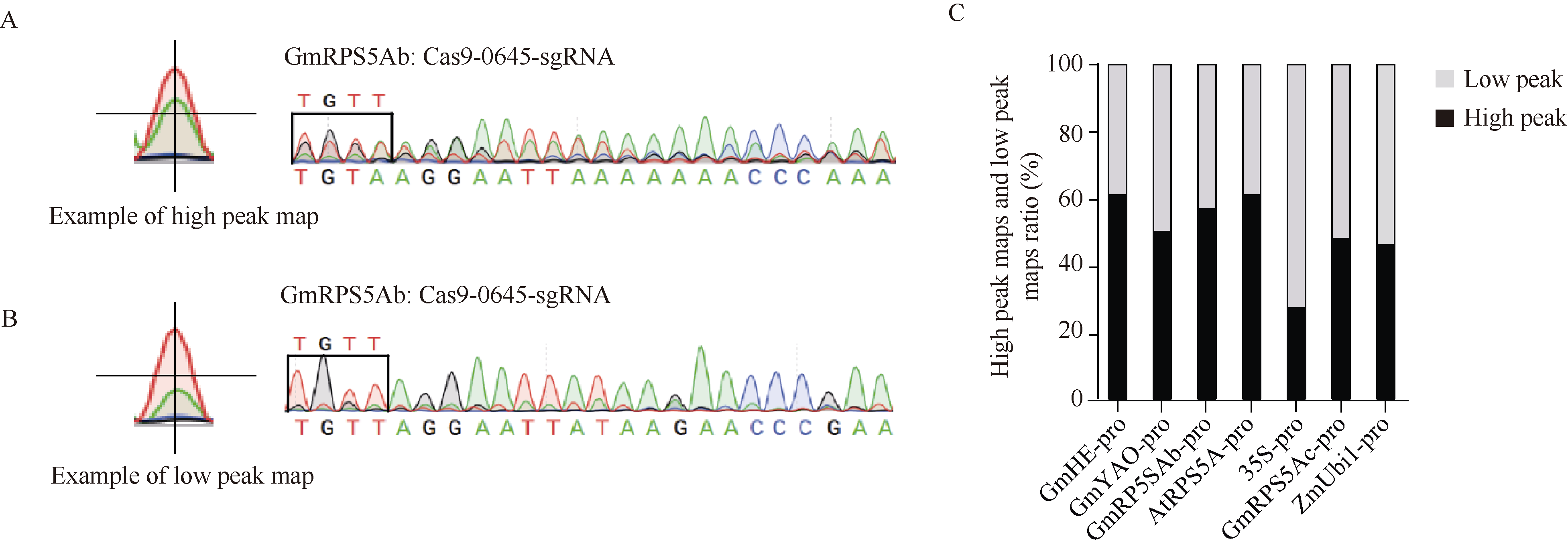
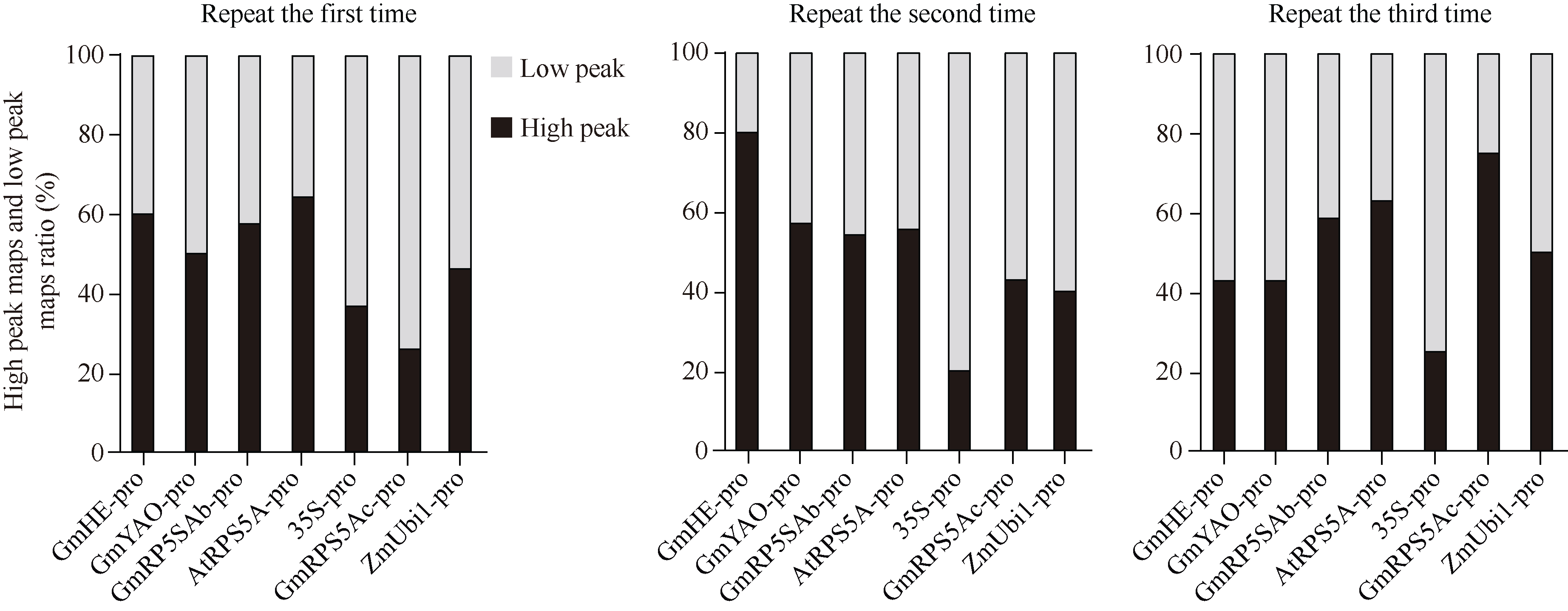
CRISPR/Cas9系统作为高效基因编辑系统, 已被广泛应用于动植物中。多种Cas9基因启动子, 如RPS5A、YAO等被报道用于提高CRISPR/Cas9系统的基因编辑效率。但在大豆中, 不同Cas9启动子对CRISPR/Cas9基因编辑系统效率的影响还没有被阐明。本研究选择了6个已知功能的高效Cas9启动子(p35S、pGmRPS5Ab、pGmRPS5Ac、pAtRPS5A、pGmYAO、pZmUbiquitin)和1个大豆内源未知功能的启动子(pGmHE), 构建CRISPR/Cas9基因敲除载体。通过农杆菌介导的大豆发根系统, 检测这些Cas9启动子对大豆内源基因GmSPA1a和GmEID1的编辑效率, 结果显示大豆内源启动子pGmRPS5Ab对靶基因的编辑效率最高, pAtRPS5A、p35S、pZmUbiquitin对下游基因的编辑效率高于pGmYAO和pGmRPS5Ac。进一步对靶位点测序峰图的分析发现, 使用了pGmRPS5Ab和pAtRPS5A启动子的测序峰图中, 高峰占比较大, 分别为64.0%和58.6%; 而使用p35S的测序峰图中, 低峰占比较高, 为63.3%。这表明pGmRP5SAb和pAtRPS5A启动子不但编辑效率高, 而且编辑效果好, 更有利于在下一代中分离出纯合突变体植株。这些结果将为构建高效大豆基因编辑载体提供参考, 为提高大豆基因编辑效率提供依据。
| [1] |
Manghwar H, Lindsey K, Zhang X, Jin S. CRISPR/Cas system: recent advances and future prospects for genome editing. Trends Plant Sci, 2019, 24: 1102-1125.
doi: S1360-1385(19)30243-2 pmid: 31727474 |
| [2] |
Christian M, Cermak T, Doyle E L, Schmidt C, Zhang F, Hummel A, Bogdanove A J, Voytas D F. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics, 2010, 186: 757-761.
doi: 10.1534/genetics.110.120717 pmid: 20660643 |
| [3] |
Wright D A, Townsend J A, Winfrey R J Jr, Irwin P A, Rajagopal J, Lonosky P M, Hall B D, Jondle M D, Voytas D F. High-frequency homologous recombination in plants mediated by zinc-finger nucleases. Plant J, 2005, 44: 693-705.
doi: 10.1111/j.1365-313X.2005.02551.x pmid: 16262717 |
| [4] |
Nekrasov V, Staskawicz B, Weigel D, Jones J D, Kamoun S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat Biotechnol, 2013, 31: 691-693.
doi: 10.1038/nbt.2655 |
| [5] |
Li J F, Norville J E, Aach J, McCormack M, Zhang D, Bush J, Church G M, Sheen J. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat Biotechnol, 2013, 31: 688-691.
doi: 10.1038/nbt.2654 |
| [6] |
Sorek R, Lawrence C M, Wiedenheft B. CRISPR-mediated adaptive immune systems in bacteria and archaea. Annu Rev Biochem, 2013, 82: 237-266.
doi: 10.1146/annurev-biochem-072911-172315 pmid: 23495939 |
| [7] |
Garneau J E, Dupuis M È, Villion M, Romero D A, Barrangou R, Boyaval P, Fremaux C, Horvath P, Magadán A H, Moineau S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature, 2010, 468: 67-71.
doi: 10.1038/nature09523 |
| [8] |
Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna J A, Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 2012, 337: 816-821.
doi: 10.1126/science.1225829 pmid: 22745249 |
| [9] |
Mohanraju P, Makarova K S, Zetsche B, Zhang F, Koonin E V, van der Oostn J. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science, 2016, 353: aad5147.
doi: 10.1126/science.aad5147 |
| [10] |
Wright A V, Nuñez J K, Doudna J A. Biology and applications of CRISPR systems: harnessing nature’s toolbox for genome engineering. Cell, 2016, 164: 29-44.
doi: 10.1016/j.cell.2015.12.035 |
| [11] |
Komor A C, Badran A H, Liu D R. CRISPR-based technologies for the manipulation of eukaryotic genomes. Cell, 2017, 168: 20-36.
doi: S0092-8674(16)31465-9 pmid: 27866654 |
| [12] |
Bao A, Burritt D J, Chen H, Zhou X, Cao D, Tran L P. The CRISPR/Cas9 system and its applications in crop genome editing. Crit Rev Biotechnol, 2019, 39: 321-336.
doi: 10.1080/07388551.2018.1554621 pmid: 30646772 |
| [13] |
Chen K, Wang Y, Zhang R, Zhang H, Gao C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu Rev Plant Biol, 2019, 70: 667-697.
doi: 10.1146/annurev-arplant-050718-100049 pmid: 30835493 |
| [14] |
Yue J J, Hong C Y, Wei P C, Tsai Y C, Lin C S. How to start your monocot CRISPR/Cas project: plasmid design, efficiency detection, and offspring analysis. Rice, 2020, 13: 9.
doi: 10.1186/s12284-019-0354-2 |
| [15] |
Zheng X, Deng W, Luo K, Duan H, Chen Y, McAvoy R, Song S, Pei Y, Li Y. The cauliflower mosaic virus (CaMV) 35S promoter sequence alters the level and patterns of activity of adjacent tissue- and organ-specific gene promoters. Plant Cell Rep, 2007, 26: 1195-1203.
pmid: 17340093 |
| [16] |
Odell J T, Nagy F, Chua N H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature, 1985, 313: 810-812.
doi: 10.1038/313810a0 |
| [17] |
Bevan M. Binary Agrobacterium vectors for plant transformation. Nucleic Acids Res, 1984, 12: 8711-8721.
doi: 10.1093/nar/12.22.8711 pmid: 6095209 |
| [18] |
Cornejo M J, Luth D, Blankenship K M, Anderson O D, Blechl A E. Activity of a maize ubiquitin promoter in transgenic rice. Plant Mol Biol, 1993, 23: 567-581.
doi: 10.1007/BF00019304 pmid: 8219091 |
| [19] |
Christensen A H, Sharrock R A, Quail P H. Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol Biol, 1992, 18: 675-689.
doi: 10.1007/BF00020010 pmid: 1313711 |
| [20] |
Ma X L, Zhang Q Y, Zhu Q L, Liu W, Chen Y, Qiu R, Wang B, Yang Z F, Li H Y, Lin Y, Xie Y Y, Shen R X, Chen S F, Wang Z, Chen Y L, Guo J X, Chen L T, Zhao X C, Dong Z C, Liu Y G. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol Plant, 2015, 8: 1274-1284.
doi: 10.1016/j.molp.2015.04.007 pmid: 25917172 |
| [21] |
Feng Z Y, Zhang B T, Ding W, Liu X D, Yang D L, Wei P L, Cao F Q, Zhu S H, Zhang F, Mao Y F, Zhu J K. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res, 2013, 23: 1229-1232.
doi: 10.1038/cr.2013.114 pmid: 23958582 |
| [22] |
Yan L H, Wei S W, Wu Y R, Hu R L, Li H J, Yang W C, Xie Q. High-efficiency genome editing in Arabidopsis using YAO promoter-driven CRISPR/Cas9 system. Mol Plant, 2015, 8: 1820-1823.
doi: 10.1016/j.molp.2015.10.004 |
| [23] |
Li H J, Liu N Y, Shi D Q, Liu J, Yang W C. YAO is a nucleolar WD40-repeat protein critical for embryogenesis and gametogenesis in Arabidopsis. BMC Plant Biol, 2010, 10: 169.
doi: 10.1186/1471-2229-10-169 |
| [24] |
Choi M, Yun J Y, Kim J H, Kim J S, Kim S T. The efficacy of CRISPR-mediated cytosine base editing with the RPS5a promoter in Arabidopsis thaliana. Sci Rep, 2021, 11: 8087.
doi: 10.1038/s41598-021-87669-y |
| [25] |
Weijers D, Franke-van Dijk M, Vencken R J, Quint A, Hooykaas P, Offringa R. An Arabidopsis minute-like phenotype caused by a semi-dominant mutation in a RIBOSOMAL PROTEIN S5gene. Development, 2001, 128: 4289-4299.
doi: 10.1242/dev.128.21.4289 pmid: 11684664 |
| [26] |
Cai Y, Chen L, Liu X, Sun S, Wu C, Jiang B, Han T, Hou W. CRISPR/Cas9-mediated genome editing in soybean hairy roots. PLoS One, 2015, 10: e0136064.
doi: 10.1371/journal.pone.0136064 |
| [27] |
Cai Y, Chen L, Sun S, Wu C, Yao W, Jiang B, Han T, Hou W. CRISPR/Cas9-mediated deletion of large genomic fragments in soybean. Int J Mol Sci, 2018, 19: 3835.
doi: 10.3390/ijms19123835 |
| [28] |
Cai Y, Chen L, Liu X, Guo C, Sun S, Wu C, Jiang B, Han T, Hou W. CRISPR/Cas9-mediated targeted mutagenesis of GmFT2a delays flowering time in soya bean. Plant Biotechnol J, 2018, 16: 176-185.
doi: 10.1111/pbi.12758 pmid: 28509421 |
| [29] |
Li C, Li Y H, Li Y, Lu H, Hong H, Tian Y, Li H, Zhao T, Zhou X, Liu J, Zhou X, Jackson S A, Liu B, Qiu L J. A domestication-associated gene GmPRR3b regulates the circadian clock and flowering time in soybean. Mol Plant, 2020, 13: 745-759.
doi: S1674-2052(20)30030-7 pmid: 32017998 |
| [30] |
Carrijo J, Illa-Berenguer E, LaFayette P, Torres N, Aragão F J L, Parrott W, Vianna G R. Two efficient CRISPR/Cas9 systems for gene editing in soybean. Transgenic Res, 2021, 30: 239-249.
doi: 10.1007/s11248-021-00246-x pmid: 33797713 |
| [31] |
Wang L, Cao C, Ma Q, Zeng Q, Wang H, Cheng Z, Zhu G, Qi J, Ma H, Nian H, Wang Y. RNA-seq analyses of multiple meristems of soybean: novel and alternative transcripts, evolutionary and functional implications. BMC Plant Biol, 2014, 14: 169.
doi: 10.1186/1471-2229-14-169 pmid: 24939556 |
| [32] | 秦超. GmPH13和GmEID1基因改良大豆纬度适应性的机制研究. 中国农业科学院博士学位论文, 北京, 2022. |
| Qin C. Mechanism Study of the GmPH13 and GmEID1 Genes in Improving the Latitudinal Adaptability of Soybean. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2022. (in Chinese with English abstract) | |
| [33] | 秦超. 大豆中GmSPAs基因的功能分析. 中国农业科学院硕士学位论文, 北京, 2018. |
| Qin C. Function Analysis of GmSPAs in Soybean. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2018. (in Chinese with English abstract) | |
| [34] |
Oh Y, Kim S G. RPS5A Promoter-driven Cas9 produces heritable virus-induced genome editing in Icotiana attenuata. Mol Cells, 2021, 44: 911-919.
doi: 10.14348/molcells.2021.0237 |
| [35] |
Tsutsui H, Higashiyama T. pKAMA-ITACHI vectors for highly efficient CRISPR/Cas9-mediated gene knockout in Arabidopsis thaliana. Plant Cell Physiol, 2017, 58: 46-56.
doi: 10.1093/pcp/pcw191 pmid: 27856772 |
| [36] |
Ye G N, Stone D, Pang S Z, Creely W, Gonzalez K, Hinchee M. Arabidopsis ovule is the target for Agrobacterium in planta vacuum infiltration transformation. Plant J, 1999, 19: 249-257.
doi: 10.1046/j.1365-313x.1999.00520.x pmid: 10476072 |
| [37] |
Maruyama D, Hamamura Y, Takeuchi H, Susaki D, Nishimaki M, Kurihara D, Kasahara R D, Higashiyama T. Independent control by each female gamete prevents the attraction of multiple pollen tubes. Dev Cell, 2013, 25: 317-323.
doi: 10.1016/j.devcel.2013.03.013 pmid: 23673333 |
| [38] |
Wang Z P, Xing H L, Dong L, Zhang H Y, Han C Y, Wang X C, Chen Q J. Egg cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genome Biol, 2015, 16: 144.
doi: 10.1186/s13059-015-0715-0 |
| [1] | 上官小霞, 杨琴莉, 李换丽. 基于CRISPR/Cas9的棉花GhbHLH71基因编辑突变体的分析[J]. 作物学报, 2024, 50(1): 138-148. |
| [2] | 杨立达, 任俊波, 彭新月, 杨雪丽, 罗凯, 陈平, 袁晓婷, 蒲甜, 雍太文, 杨文钰. 施氮与种间距离下大豆/玉米带状套作作物生长特性及其对产量形成的影响[J]. 作物学报, 2024, 50(1): 251-264. |
| [3] | 石宇欣, 刘欣玥, 孙建强, 李晓菲, 郭潇阳, 周雅, 邱丽娟. 利用CRISPR-CAS9技术编辑GmBADH1基因改变大豆耐盐性[J]. 作物学报, 2024, 50(1): 100-109. |
| [4] | 袁晓婷, 王甜, 罗凯, 刘姗姗, 彭新月, 杨立达, 蒲甜, 王小春, 杨文钰, 雍太文. 带宽和株距对带状间作大豆物质积累分配及产量形成的影响[J]. 作物学报, 2024, 50(1): 161-171. |
| [5] | 胡艳娟, 薛丹, 耿嫡, 朱末, 王天穹, 王晓雪. 水稻OsCDF1基因突变效应及其基因组变异分析[J]. 作物学报, 2023, 49(9): 2362-2372. |
| [6] | 左春阳, 李亚玮, 李焱龙, 金双侠, 朱龙付, 张献龙, 闵玲. 陆地棉漆酶基因家族成员表达模式分析[J]. 作物学报, 2023, 49(9): 2344-2361. |
| [7] | 韦新宇, 曾跃辉, 杨旺兴, 肖长春, 候新坡, 黄建鸿, 邹文广, 许旭明. 利用CRISPR-Cas9技术编辑Badh2基因创制优质香型籼稻三系不育系[J]. 作物学报, 2023, 49(8): 2144-2159. |
| [8] | 李刚, 周彦辰, 熊亚俊, 陈伊洁, 郭庆元, 高杰, 宋健, 王俊, 李英慧, 邱丽娟. 大豆叶型调控基因Ln及其同源基因单倍型分析[J]. 作物学报, 2023, 49(8): 2051-2063. |
| [9] | 刘亭萱, 谷勇哲, 张之昊, 王俊, 孙君明, 邱丽娟. 基于高密度遗传图谱定位大豆蛋白质含量相关的QTL[J]. 作物学报, 2023, 49(6): 1532-1541. |
| [10] | 李慧, 路依萍, 汪小凯, 王璐瑶, 邱婷婷, 张雪婷, 黄海燕, 崔晓玉. CBL互作蛋白激酶GmCIPK10增强大豆耐盐性[J]. 作物学报, 2023, 49(5): 1272-1281. |
| [11] | 雷建峰, 李月, 代培红, 赵燚, 尤扬子, 贾建国, 赵帅, 曲延英, 刘晓东. 棉花中不同植物病毒介导的VIGE体系的研究[J]. 作物学报, 2023, 49(4): 978-987. |
| [12] | 吴宗声, 徐彩龙, 李瑞东, 徐一帆, 孙石, 韩天富, 宋雯雯, 吴存祥. 麦秸覆盖还田对大豆耕层物理性状及产量形成的影响[J]. 作物学报, 2023, 49(4): 1052-1064. |
| [13] | 舒泽兵, 罗万宇, 蒲甜, 陈国鹏, 梁冰, 杨文钰, 王小春. 基于高产与高效条件下鲜食玉米鲜食大豆带状间作田间配置技术优化[J]. 作物学报, 2023, 49(4): 1140-1150. |
| [14] | 刘姗姗, 庞婷, 袁晓婷, 罗凯, 陈平, 付智丹, 王小春, 杨峰, 雍太文, 杨文钰. 种间距对不同结瘤特性套作大豆根瘤生长及固氮潜力的影响[J]. 作物学报, 2023, 49(3): 833-844. |
| [15] | 杨硕, 武阳春, 刘鑫磊, 唐晓飞, 薛永国, 曹旦, 王婉, 刘亭萱, 祁航, 栾晓燕, 邱丽娟. 大豆蛋白含量主效位点qPRO-20-1的精细定位[J]. 作物学报, 2023, 49(2): 310-320. |
|