欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (6): 1562-1572.doi: 10.3724/SP.J.1006.2023.23042
马娟*( ), 朱卫红, 刘京宝, 宇婷, 黄璐, 郭国俊
), 朱卫红, 刘京宝, 宇婷, 黄璐, 郭国俊
MA Juan*(), ZHU Wei-Hong, LIU Jing-Bao, YU Ting, HUANG Lu, GUO Guo-Jun
摘要:
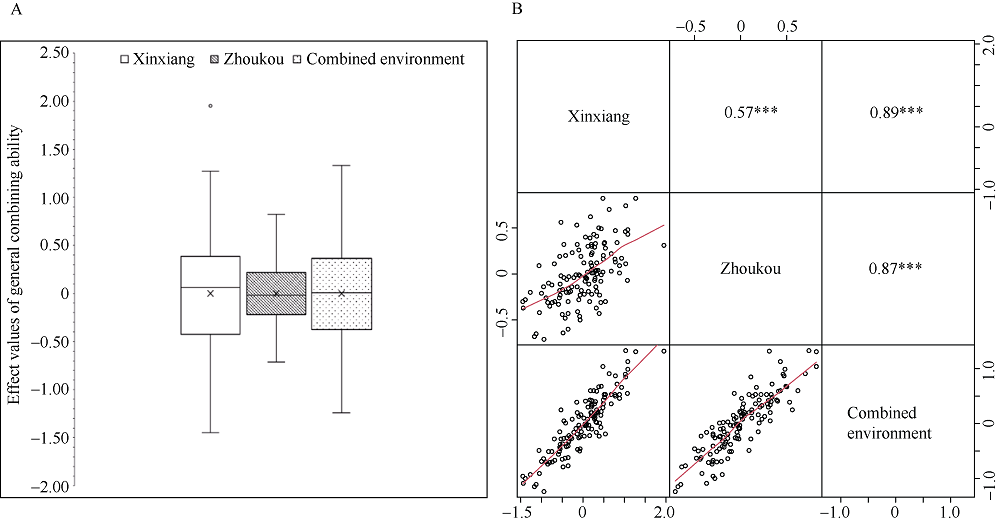
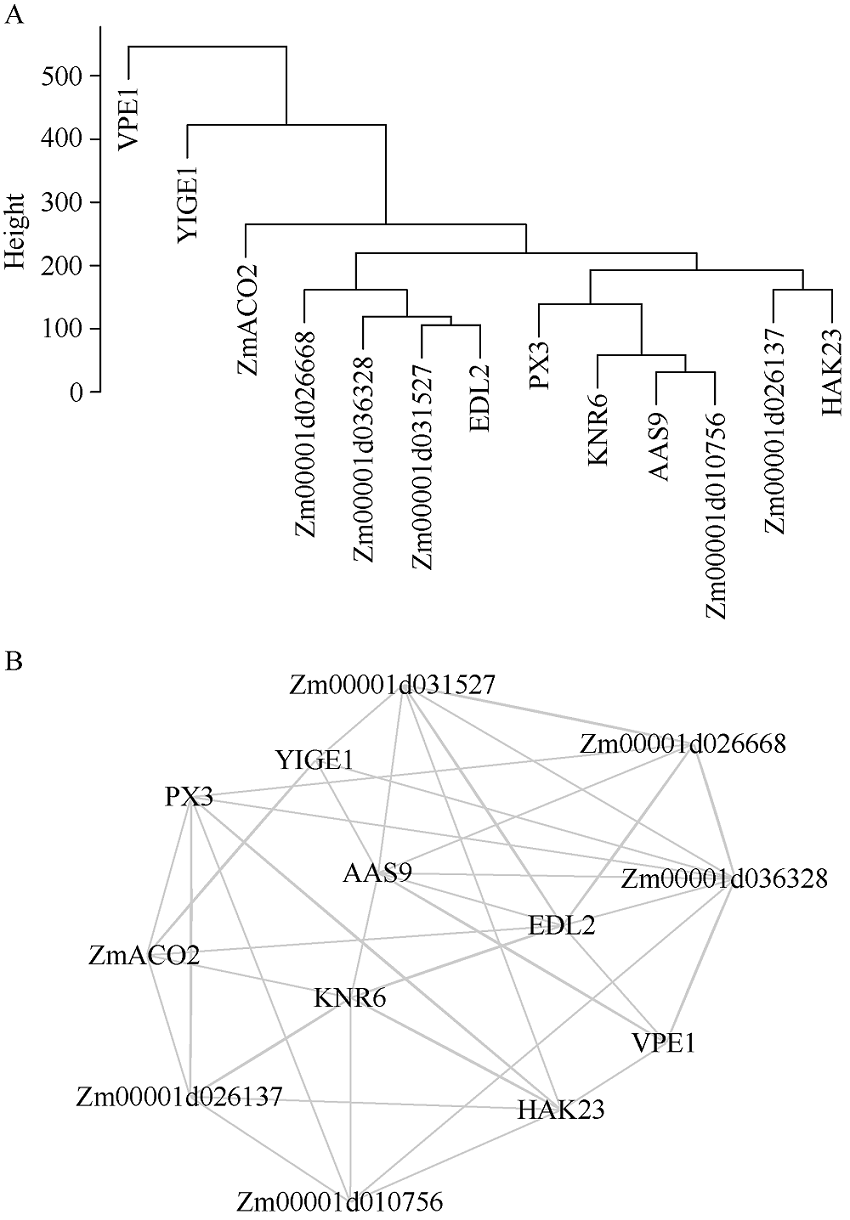
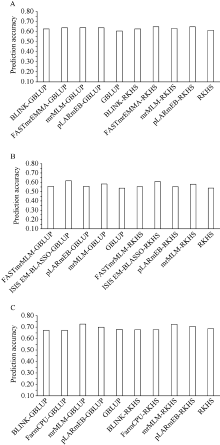
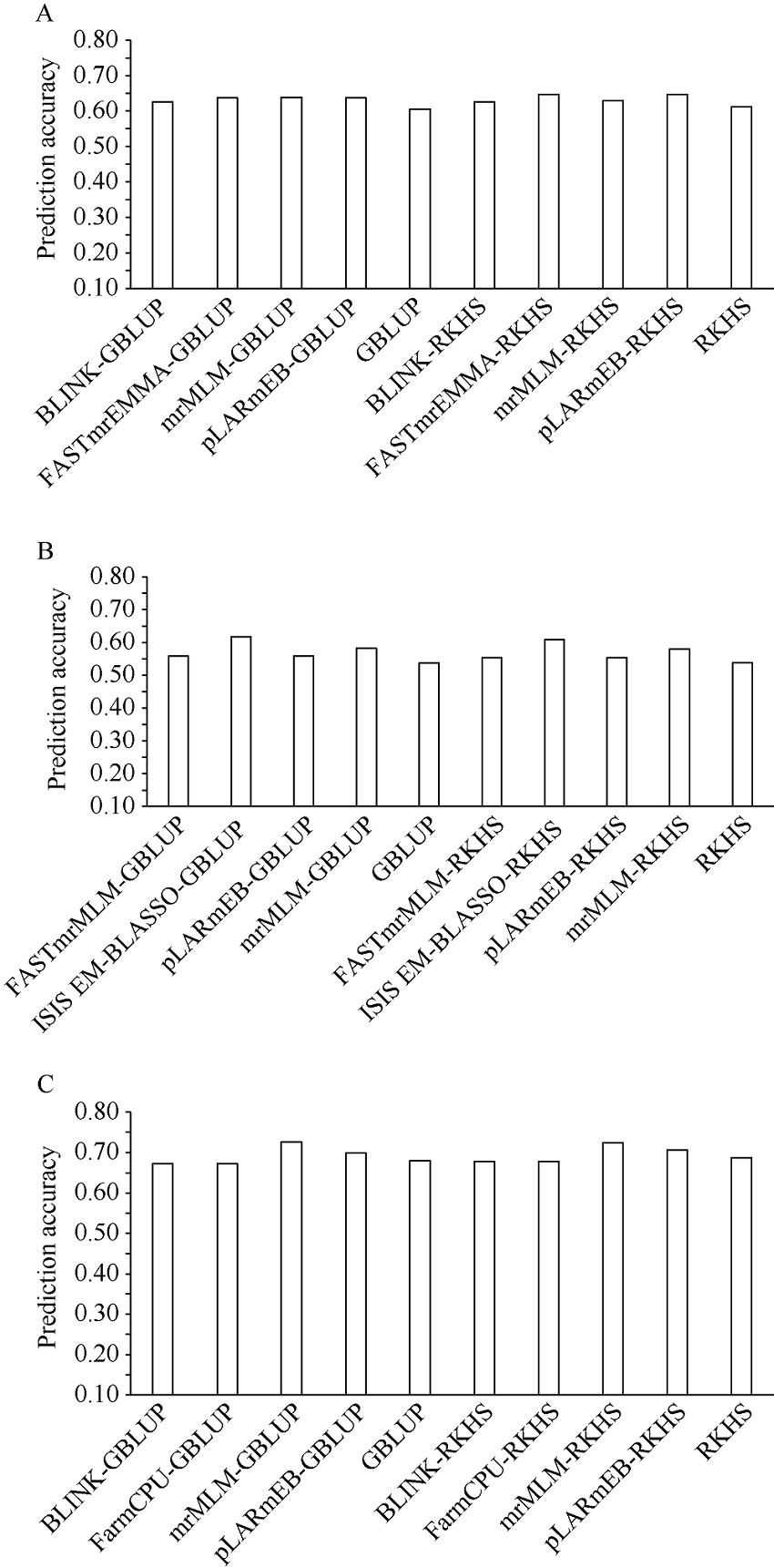
穗长是一个重要的农艺性状, 与产量密切相关。一般配合力(general combining ability, GCA)是评价优异自交系的重要指标。因此, 解析穗长GCA的遗传基础, 制定相应的育种策略对玉米杂交种产量的提高具有重要意义。本研究以123个玉米自交系和8个测验种按照North Carolina II遗传交配设计组配的537个F1杂交种为试验材料, 在2个环境下进行表型鉴定, 利用玉米5.5 K液相育种芯片鉴定的11,734个SNP (single nucleotide polymorphisms)对2个环境以及综合环境穗长GCA进行多位点全基因组关联分析(multi-locus genome-wide association study, MGWAS)和基因组预测。利用7种MGWAS共检测到11个穗长GCA显著关联SNP标记(P < 8.52E-07), 单个位点解释GCA变异介于8.06%~28.23%之间。不同MGWAS共定位的SNP位点有5个。位点7_178103602在周口和综合环境利用mrMLM (multi-locus random-SNP-effect mixed linear model)方法重复检测到, 可解释穗长GCA变异的26.02%~28.23%, 为环境稳定的主效SNP。共挖掘10个候选基因, 其中auxin amido synthetase 9和EID1-like F-box protein 2可能是控制穗长GCA的关键基因。5种随机效应模型对3个环境穗长GCA的预测准确性介于0.53~0.69之间, 且模型间差异较小。在新乡和周口环境, GBLUP (genomic best linear unbiased prediction)和RKHS (reproducing kernel Hilbert space)整合不同显著位点作为固定效应均可提高穗长GCA基因组估计育种值的准确性, 提高率为2.34%~14.98%, 而在综合环境中除了利用FarmCPU (fixed and random model circulating probability unification)或BLINK (Bayesian-information and linkage-disequilibrium iteratively nested keyway)鉴定的1个显著位点作为固定效应会略降低预测精度外, 其他2种MGWAS方法显著位点的加入均能提高基因组预测力, 提高率为2.80%~6.84%。因此, MGWAS显著位点作为固定效应加入预测模型有利于提高穗长GCA基因组估计育种值的准确性, 可用来对玉米亲本穗长GCA进行有效预测和选择。
| [1] | 张人予. 玉米穗长基因EL3的克隆及我国优良自交系基因组变异分析. 中国农业大学博士学位论文, 北京, 2018. |
| Zhang R Y. Cloning of EL3 for Ear Length in Maize and Patterns of Genomic Variation in Chinese Maize Inbred Lines. PhD Dissertation of China Agricultural University, Beijing, China, 2018. (in Chinese with English abstract) | |
| [2] |
Jia H T, Li M F, Li W Y, Liu L, Jian Y N, Yang Z X, Shen X M, Ning Q, Du Y F, Zhao R, Jackson D, Yang X H, Zhang Z X. A serine/threonine protein kinase encoding gene KERNEL NUMBER PER ROW6 regulates maize grain yield. Nat Commun, 2020, 11: 988-998.
doi: 10.1038/s41467-020-14746-7 |
| [3] | 周广飞. 一个控制玉米行粒数、穗长其一般配合力的多效性QTL (qKNR7.2)鉴定. 华中农业大学硕士学位论文, 湖北武汉, 2014. |
| Zhou G F. Identification of A Pleitropic QTL (qKNR7.2) for Kernel Row Number Per Row, Ear Length, and General Combining Ability of Maize. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2014. (in Chinese with English abstract) | |
| [4] |
Liu X G, Hu X X, Li K, Liu Z F, Wu Y J, Feng G, Huang C L, Wang H W. Identifying quantitative trait loci for the general combining ability of yield-relevant traits in maize. Breed Sci, 2021, 71: 217-228.
doi: 10.1270/jsbbs.20008 |
| [5] | 监立强. 玉米产量相关性状及其一般配合力的关联分析. 河北农业大学硕士学位论文, 河北保定, 2017. |
| Jian L Q. Genome-Wide Association Study of Yield-Related Traits and General Combining Ability in Maize (Zea mays L.). MS Thesis of Hebei Agricultural University, Baoding, Hebei, China, 2017. (in Chinese with English abstract) | |
| [6] | 刘文童, 监立强, 郭晋杰, 赵永锋, 黄亚群, 陈景堂, 祝丽英. 玉米穗部性状及其一般配合力的关联分析. 植物遗传资源学报, 2020, 21: 706-715. |
| Liu W T, Jian L Q, Guo J J, Zhao Y F, Huang Y Q, Chen J C, Zhu L Y. Association analysis of ear-related traits and their general combining ability in maize. J Plant Genetic Resour, 2020, 21: 706-715. (in Chinese with English abstract) | |
| [7] | 温阳俊, 冯建英, 张瑾. 多位点关联分析方法学的研究进展. 南京农业大学学报, 2022, 45: 1-10. |
| Wen Y J, Feng J, Zhang J. Research progress of mulit-locus genome-wide association study. J Nanjing Agric Univ, 2022, 45: 1-10. (in Chinese with English abstract) | |
| [8] | Liu X L, Huang M, Fan B, Buckler E S, Zhang Z. Iterative usage of fixed and random effect models for powerful and efficient genome wide association studies. PLoS Genet, 2016, 12: e1005767. |
| [9] | Huang M, Liu X, Zhou Y, Summers R M, Zhang Z W. BLINK: a package for the next level of genome-wide association studies with both individuals and markers in the millions. Gigascience, 2019, 8: 1-12. |
| [10] |
Wang S B, Feng J Y, Ren W L, Huang B, Zhou L, Wen Y J, Zhang J, Dunwell J M, Xu S, Zhang Y M. Improving power and accuracy of genome-wide association studies via a multi-locus mixed linear model methodology. Sci Rep, 2016, 6: 19444-19453.
doi: 10.1038/srep19444 |
| [11] | Tamba C L, Zhang Y M. A fast mrMLM algorithm for multi-locus genome-wide association studies. BioRxiv, 2018. https://doi.org/10.1101/341784. |
| [12] |
Wen Y J, Zhang H, Ni Y L, Huang B, Zhang J, Feng J Y, Wang S B, Dunwell J M, Zhang Y M, Wu R. Methodological implementation of mixed linear models in multi-locus genome-wide association studies. Brief Bioinform, 2018, 19: 700-712.
doi: 10.1093/bib/bbw145 |
| [13] |
Zhang J, Feng J Y, Ni Y L, Wen Y J, Niu Y, Tamba C L, Yue C, Song Q, Zhang Y M. pLARmEB: integration of least angle regression with empirical Bayes for multilocus genome-wide association studies. Heredity, 2017, 118: 517-524.
doi: 10.1038/hdy.2017.8 pmid: 28295030 |
| [14] | Tamba C L, Ni Y L, Zhang Y M. Iterative sure independence screening EM-Bayesian LASSO algorithm for multi-locus genome-wide association studies. PLoS Comput Biol, 2017, 13: e1005357. |
| [15] |
Yang Y, Chai Y M, Zhang X, Lu S, Zhao Z C, Wei D, Chen L, Hu Y G. Multi-locus GWAS of quality traits in bread wheat: mining more candidate genes and possible regulatory network. Front Plant Sci, 2020, 11: 1091-1109.
doi: 10.3389/fpls.2020.01091 pmid: 32849679 |
| [16] |
Peng Y C, Liu H B, Chen J, Shi T T, Zhang C, Sun D F, He Z H, Hao Y F, Chen W. Genome-wide association studies of free amino acid levels by six multi-locus models in bread wheat. Front Plant Sci, 2018, 9: 1196-1204.
doi: 10.3389/fpls.2018.01196 pmid: 30154817 |
| [17] |
Su J J, Wang C X, Hao F S, Ma Q, Wang J, Li J L, Ning X Z. Genetic detection of lint percentage applying single-locus and multi-locus genome-wide association studies in Chinese early-maturity upland cotton. Front Plant Sci, 2019, 10: 964-974.
doi: 10.3389/fpls.2019.00964 pmid: 31428110 |
| [18] |
Cui Y R, Zhang F, Zhou Y L. The application of multi-locus GWAS for the detection of salt-tolerance loci in rice. Front Plant Sci, 2018, 9: 1464-1472.
doi: 10.3389/fpls.2018.01464 pmid: 30337936 |
| [19] |
Zhou G F, Zhu Q L, Mao Y X, Chen G Q, Xue L, Lu H H, Shi M L, Zhang Z L, Song X D, Zhang H M, Hao D R. Multi-locus genome-wide association study and genomic selection of kernel moisture content at the harvest stage in maize. Front Plant Sci, 2021, 12: 697688-697700.
doi: 10.3389/fpls.2021.697688 |
| [20] |
Meuwissen T H, Hayes B J, Goddard M E. Prediction of total genetic value using genome-wide dense marker maps. Genetics, 2001, 157: 1819-1829.
doi: 10.1093/genetics/157.4.1819 pmid: 11290733 |
| [21] |
Vanraden P M. Efficient methods to compute genomic predictions. J Dairy Sci, 2008, 91: 4414-4423.
doi: 10.3168/jds.2007-0980 pmid: 18946147 |
| [22] |
de los Campos G, Naya H, Gianola D, Crossa J, Legarra A, Manfredi E, Weigel K, Cotes J M. Predicting quantitative traits with regression models for dense molecular markers and pedigree. Genetics, 2009, 182: 375-385.
doi: 10.1534/genetics.109.101501 pmid: 19293140 |
| [23] |
González-Recio O, Forni S. Genome-wide prediction of discrete traits using bayesian regressions and machine learning. Genet Sel Evol, 2011, 43: 7-18.
doi: 10.1186/1297-9686-43-7 pmid: 21329522 |
| [24] |
Guo Z G, Tucker D M, Lu J, Kishore V, Gay G. Evaluation of genome-wide selection efficiency in maize nested association mapping populations. Theor Appl Genet, 2012, 124: 261-275.
doi: 10.1007/s00122-011-1702-9 pmid: 21938474 |
| [25] |
Lian L, Jacobson A, Zhong S Q. Genome wide prediction accuracy within 969 maize biparental populations. Crop Sci, 2014, 54: 1514-1522.
doi: 10.2135/cropsci2013.12.0856 |
| [26] |
Technow F, Schrag T A, Schipprack W, Bauer E, Simianer H, Melchinger A E. Genome properties and prospects of genomic prediction of hybrid performance in a breeding program of maize. Genetics, 2014, 197: 1343-1355.
doi: 10.1534/genetics.114.165860 pmid: 24850820 |
| [27] |
de Oliveira A A, Resende M F R Jr, Ferrão L F V, Amadeu R R, Guimarães L J M, Guimarães C T, Pastina M M, Margarido G R A. Genomic prediction applied to multiple traits and environments in second season maize hybrids. Heredity, 2020, 125: 60-72.
doi: 10.1038/s41437-020-0321-0 pmid: 32472060 |
| [28] |
Wang X, Zhang Z L, Xu Y, Li P P, Xu C W. Using genomic data to improve the estimation of general combining ability based on sparse partial diallel cross designs in maize. Crop J, 2020, 8: 819-829.
doi: 10.1016/j.cj.2020.04.012 |
| [29] |
Zhang A, Pérez-Rodríguez P, Vicente F S, Palacios-Rojas N, Dhliwayo T, Liu Y B, Cui Z H, Guan Y, Wang H, Zheng H J, Olsen M, Prasanna B M, Ruan Y Y, Crossa J, Zhang X C. Genomic prediction of the performance of hybrids and the combining abilities for line by tester trials in maize. Crop J, 2021, 10: 109-116.
doi: 10.1016/j.cj.2021.04.007 |
| [30] | 熊雪航, 段海洋, 李文龙, 李建新, 孙莉, 孙岩, 秦永田, 汤继华, 张雪海. 玉米穗长全基因组关联分析. 分子植物育种, 2022, https://kns.cnki.net/kcms/detail/46.1068.S.20220630.1359.004.html |
| Xiong X H, Duan H Y, Li W L, Li J X, Sun L, Sun Y, Qin Y T, Tang J H, Zhang X H. Genome-wide association study of ear length in maize. Mol Plant Breed, 2022, https://kns.cnki.net/kcms/detail/46.1068.S.20220630.1359.004.html. (in Chinese with English abstract) | |
| [31] |
秦文萱, 鲍建喜, 王彦博, 马雅杰, 龙艳, 李金萍, 董振营, 万向元. 玉米叶夹角性状的全基因组关联分析与关键位点优异等位变异挖掘. 作物学报, 2022, 48: 2691-2705.
doi: 10.3724/SP.J.1006.2022.23019 |
| Qin W X, Bao J X, Wang Y B, Ma Y J, Long Y, Li J P, Dong Z Y, Wang X Y. Genome-wide association study of leaf angle traits and mining of elite alleles from the major loci in maize. Acta Agron Sin, 2022, 48: 2691-2705 (in Chinese with English abstract). | |
| [32] |
彭勃, 赵晓雷, 王奕, 袁文娅, 李春辉, 李永祥, 张登峰, 石云素, 宋燕春, 王天宇, 黎裕. 玉米叶向值的全基因组关联分析. 作物学报, 2020, 46: 819-831.
doi: 10.3724/SP.J.1006.2020.93063 |
|
Peng B, Zhao X L, Wang Y, Yuan Y W, Li C H, Li Y X, Zhang D F, Shi S Y, Song C Y, Wang T Y, Li Y. Genome-wide association studies of leaf orientation value in maize. Acta Agron Sin, 2020, 46: 819-831. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2020.93063 |
|
| [33] |
Pritchard J K, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics, 2000, 155: 945-959.
doi: 10.1093/genetics/155.2.945 pmid: 10835412 |
| [34] |
Jakobsson M, Rosenberg N A. CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics, 2007, 23: 1801-1806.
doi: 10.1093/bioinformatics/btm233 pmid: 17485429 |
| [35] |
Bradbury P J, Zhang Z W, Kroon D E, Casstevens T M, Ramdoss Y, Buckler E S. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635.
doi: 10.1093/bioinformatics/btm308 pmid: 17586829 |
| [36] |
Wang J B, Zhang Z W. GAPIT Version 3: boosting power and accuracy for genomic association and prediction. Genom Proteom Bioinf, 2021, 19: 629-640.
doi: 10.1016/j.gpb.2021.08.005 pmid: 34492338 |
| [37] |
Zhang Y W, Tamba C L, Wen Y J, Li P, Ren W L, Ni Y L, Gao J, Zhang Y M. mrMLM v4.0.2: an R platform for multi-locus genome-wide association studies. Genom Proteom Bioinf, 2020, 18: 481-487.
doi: 10.1016/j.gpb.2020.06.006 |
| [38] |
Cingolani P, Platts A, Wang L, Coon M, Nguyen T, Wang L, Land S J, Lu X, Ruden D M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly (Austin), 2012, 6: 80-92.
doi: 10.4161/fly.19695 pmid: 22728672 |
| [39] |
Luo Y, Zhang M L, Liu Y, Liu J, Li W Q, Chen G S, Peng Y, Jin M, Wei W, Jian L, Yan J, Fernie A R, Yan J B. Genetic variation in YIGE1 contributes to ear length and grain yield in maize. New Phytol, 2022, 234: 513-526.
doi: 10.1111/nph.v234.2 |
| [40] |
Ning Q, Jian Y N, Du Y, Li Y F, Shen X M, Jia H T, Zhao R, Zhan J M, Yang F, Jackson D, Liu L, Zhang Z W. An ethylene biosynthesis enzyme controls quantitative variation in maize ear length and kernel yield. Nat Commun, 2021, 12: 5832-5842.
doi: 10.1038/s41467-021-26123-z pmid: 34611160 |
| [41] |
Pérez P, de los Campos G. Genome-wide regression and prediction with the BGLR statistical package. Genetics, 2014, 198: 483-495.
doi: 10.1534/genetics.114.164442 pmid: 25009151 |
| [42] |
Zhang Y M, Jia Z, Dunwell J M. The applications of new multi-locus GWAS methodologies in the genetic dissection of complex traits. Front Plant Sci, 2019, 10: 100-105.
doi: 10.3389/fpls.2019.00100 |
| [43] |
An Y X, Chen L, Li Y X, Li C H, Shi Y S, Zhang D F, Li Y, Wang T Y. Genome-wide association studies and whole-genome prediction reveal the genetic architecture of KRN in maize. BMC Plant Biol, 2020, 20: 490-500.
doi: 10.1186/s12870-020-02676-x pmid: 33109077 |
| [44] | Zhou B, Zhou Z J, Ding J Q, Zhang X C, Mu C, Wu Y, Gao J Y, Song Y X, Wang S W, Ma J L, Li X T, Wang R X, Xia Z L, Chen J F, Wu J Y. Combining three mapping strategies to reveal quantitative trait loci and candidate genes for maize ear length. Plant Genome, 2018, 11: 1-8. |
| [45] |
Li D D, Zhou Z Q, Lu X H, Jiang Y, Li G L, Li J H, Wang H Y, Chen S J, Li X H, Würschum T, Reif J C, Xu S Z, Li M S, Liu W X. Genetic dissection of hybrid performance and heterosis for yield-related traits in maize. Front Plant Sci, 2021, 12: 774478-774496.
doi: 10.3389/fpls.2021.774478 |
| [46] |
Su C F, Wang W, Gong S L, Zuo J H, Li S J, Xu S Z. High density linkage map construction and mapping of yield trait QTLs in maize (Zea mays) using the genotyping-by-sequencing (GBS) technology. Front Plant Sci, 2017, 8: 706-719.
doi: 10.3389/fpls.2017.00706 |
| [47] |
Chen L, An Y X, Li Y X, Li C H, Shi Y S, Song Y C, Zhang D F, Wang T Y, Li Y. Candidate loci for yield-related traits in maize revealed by a combination of MetaQTL analysis and regional association mapping. Front Plant Sci, 2017, 8: 2190-2203.
doi: 10.3389/fpls.2017.02190 pmid: 29312420 |
| [48] |
Zhou Z P, Li G L, Tan S Y, Li D D, Liu W X. A QTL atlas for grain yield and its component traits in maize (Zea mays). Plant Breed, 2020, 139: 562-574.
doi: 10.1111/pbr.v139.3 |
| [49] |
Zhao Y M, Su C F. Mapping quantitative trait loci for yield-related traits and predicting candidate genes for grain weight in maize. Sci Rep, 2019, 9: 16112-16121.
doi: 10.1038/s41598-019-52222-5 pmid: 31695075 |
| [50] |
Lu X, Zhou Z Q, Yuan Z H, Zhang C S, Hao Z F, Wang Z H, Li M S, Zhang D G, Yong H J, Han J N, Li X H, Weng J F. Genetic dissection of the general combining ability of yield-related traits in maize. Front Plant Sci, 2020, 11: 788-802.
doi: 10.3389/fpls.2020.00788 pmid: 32793248 |
| [51] |
Galli M, Liu Q J, Moss B L, Malcomber S, Li W, Gaines C, Federici S, Roshkovan J, Meeley R, Nemhauser J L, Gallavotti A. Auxin signaling modules regulate maize inflorescence architecture. Proc Natl Acad Sci USA, 2015, 112: 13372-13377.
doi: 10.1073/pnas.1516473112 pmid: 26464512 |
| [52] |
Koops P, Pelser S, Ignatz M, Klose C, Marrocco-Selden K, Kretsch T. EDL3 is an F-box protein involved in the regulation of abscisic acid signalling in Arabidopsis thaliana. J Exp Bot, 2011, 62: 5547-5560.
doi: 10.1093/jxb/err236 |
| [53] |
Zhang H H, Yin L L, Wang M Y, Yuan X H, Liu X L. Factors affecting the accuracy of genomic selection for agricultural economic traits in maize, cattle, and pig populations. Front Genet, 2019, 10: 189-198.
doi: 10.3389/fgene.2019.00189 pmid: 30923535 |
| [54] |
Tehseen M M, Kehel Z, Sansaloni C P, Lopes M D S, Amri A, Kurtulus E, Nazari K. Comparison of genomic prediction methods for yellow, stem, and leaf rust resistance in wheat landraces from Afghanistan. Plants, 2021, 10: 558-572.
doi: 10.3390/plants10030558 |
| [55] |
Ma J, Cao Y Y. Genetic dissection of grain yield of maize and yield-related traits through association mapping and genomic prediction. Front Plant Sci, 2021, 12: 690059-690069.
doi: 10.3389/fpls.2021.690059 |
| [56] |
Lozada D N, Mason R E, Sarinelli J M, Brown-Guedira G. Accuracy of genomic selection for grain yield and agronomic traits in soft red winter wheat. BMC Genet, 2019, 20: 82.
doi: 10.1186/s12863-019-0785-1 pmid: 31675927 |
| [57] |
Odilbekov F, Armoniené R, Koc A, Svensson J, Chawade A. GWAS-assisted genomic prediction to predict resistance to Septoria tritici Blotch in Nordic winter wheat at seedling stage. Front Genet, 2019, 10: 1224-1233.
doi: 10.3389/fgene.2019.01224 pmid: 31850073 |
| [58] | 马娟, 朱卫红, 丁俊强. 玉米重要农艺性状的基因组预测分析. 玉米科学, 2022, 30(1): 48-52. |
| Ma J, Zhu W H, Ding J Q. Genomic prediction analysis for maize important agronomic traits. J Maize Sci, 2022, 30(1): 48-52. (in Chinese with English abstract) | |
| [59] |
Arruda M, Lipka A, Brown P, Krill A, Thurber C, Brown-Guedira G, Dong Y, Foresman B J, Kolb F L. Comparing genomic selection and marker-assisted selection for Fusarium head blight resistance in wheat (Triticum aestivum). Mol Breed, 2016, 36: 1-11.
doi: 10.1007/s11032-015-0425-z |
| [60] |
Bernardo R. Genomewide selection when major genes are known. Crop Sci, 2014, 54: 68-75.
doi: 10.2135/cropsci2013.05.0315 |
| [61] |
Yuan Y, Cairns J E, Babu R, Gowda M, Makumbi D, Magorokosho C, Zhang A, Liu Y B, Wang N, Hao Z F, San V F, Olsen M S, Prasanna B M, Lu Y L, Zhang X C. Genome-wide association mapping and genomic prediction analyses reveal the genetic architecture of grain yield and flowering time under drought and heat stress conditions in maize. Front Plant Sci, 2019, 9: 1919-1933.
doi: 10.3389/fpls.2018.01919 |
| [62] |
Liu Y B, Hu G H, Zhang A, Loladze A, Hu Y X, Wang H, Qu J T, Zhang X C, Olsen M, Vicente F S, Crossa J, Lin F, Prasanna B M. Genome-wide association study and genomic prediction of Fusarium ear rot resistance in tropical maize germplasm. Crop J, 2021, 9: 325-341.
doi: 10.1016/j.cj.2020.08.008 |
| [63] | Cericola F, Jahoor A, Orabi J, Andersen J R, Janss L L, Jensen J. Optimizing training population size and genotyping strategy for genomic prediction using association study results and pedigree information: a case of study in advanced wheat breeding lines. PLoS One, 2017, 12: e0169606. |
| [1] | 许加波, 吴鹏昊, 黄博文, 陈占辉, 马月虹, 任姣姣. 利用F2:3家系来源单倍体定位玉米雄穗相关性状QTL及全基因组选择[J]. 作物学报, 2023, 49(3): 622-633. |
| [2] | 严威凯. 品种选育与评价的原理和方法评述[J]. 作物学报, 2022, 48(9): 2137-2154. |
| [3] | 贺建波,刘方东,邢光南,王吴彬,赵团结,管荣展,盖钧镒. 限制性两阶段多位点全基因组关联分析方法的特点与计算程序[J]. 作物学报, 2018, 44(9): 1274-1289. |
| [4] | 马岩松,刘章雄,文自翔,魏淑红,杨春明,王会才,杨春燕,卢为国,徐冉,张万海,吴纪安,胡国华,栾晓燕,付亚书,郭. 群体构成方式对大豆百粒重全基因组选择预测准确度的影响[J]. 作物学报, 2018, 44(01): 43-52. |
| [5] | 王博新,王亚辉,陈朋飞,刘徐冬雨,冯志前,郝引川,张仁和,张兴华,薛吉全*. 源于陕A群、陕B群玉米自交系在不同密度条件下配合力分析[J]. 作物学报, 2017, 43(09): 1328-1336. |
| [6] | 王建军,雍洪军,张晓聪,李明顺,张德贵,白丽,高志强,张世煌,李新海. 12个外来玉米群体与我国主要种质配合力效应和杂种优势分析[J]. 作物学报, 2012, 38(12): 2170-2177. |
| [7] | 束永俊, 吴磊, 王丹, 郭长虹. 人工神经网络在作物基因组选择中的应用[J]. 作物学报, 2011, 37(12): 2179-2186. |
| [8] | 陈洪梅, 汪燕芬, 姚文华, 罗黎明, 李佳莉, 徐春霞, 番兴明, 郭华春. 导入热带种质的温带玉米自交系的利用潜力[J]. 作物学报, 2011, 37(10): 1785-1793. |
| [9] | 杨加银,盖钧镒. 黄淮地区大豆重要亲本间产量的杂种优势、配合力及其遗传基础[J]. 作物学报, 2009, 35(4): 620-630. |
|