欢迎访问作物学报,今天是
作物学报 ›› 2018, Vol. 44 ›› Issue (05): 672-685.doi: 10.3724/SP.J.1006.2018.00672
田润苗1,*( ), 张雪海1,*, 汤继华1, 白光红2, 付志远1,*()
), 张雪海1,*, 汤继华1, 白光红2, 付志远1,*()
Run-Miao TIAN1,**(), Xue-Hai ZHANG1,**, Ji-Hua TANG1, Guang-Hong BAI2, Zhi-Yuan FU1,*()
摘要:
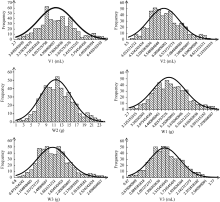
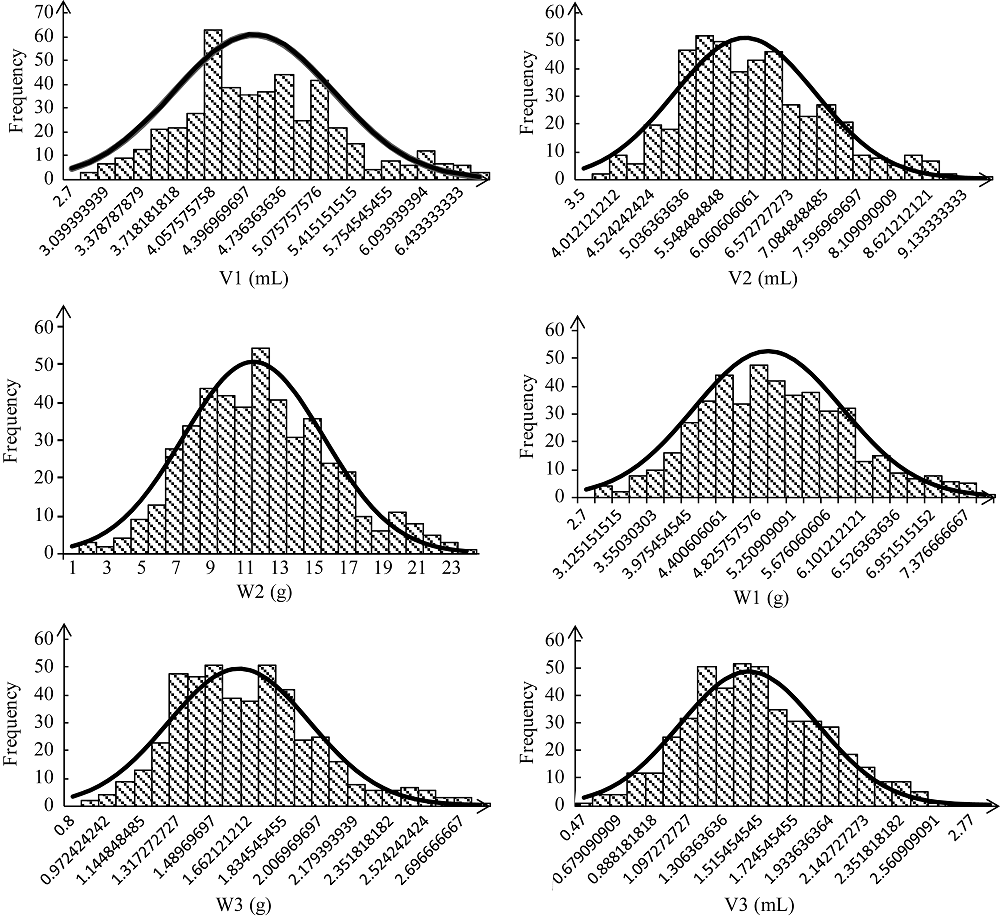
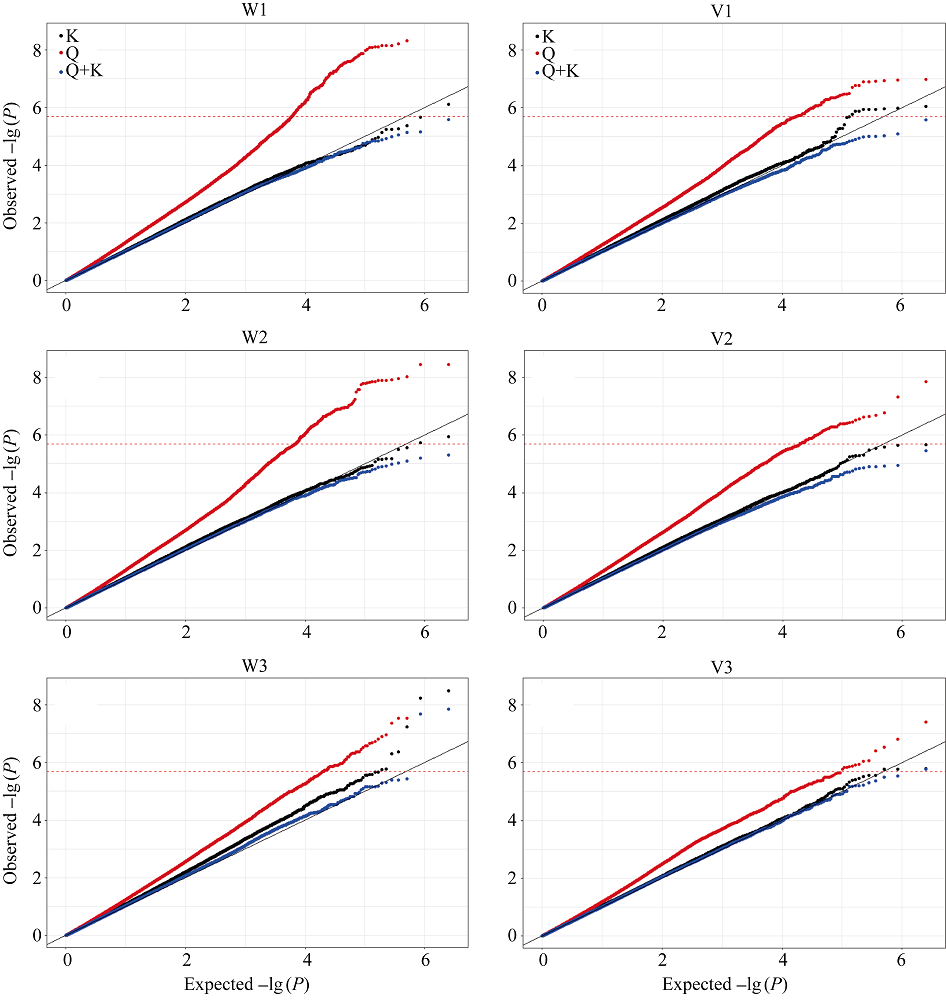
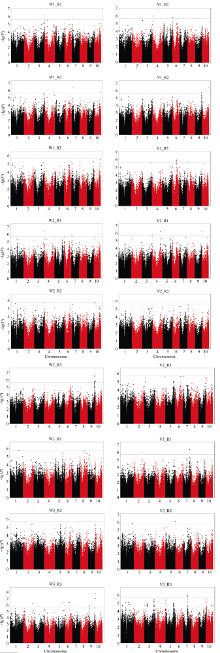
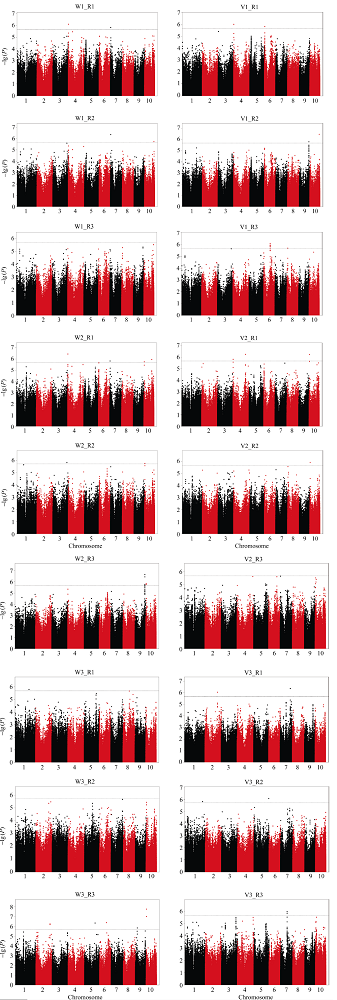
种子萌发是出苗的前提, 对玉米产量影响重大。为了解玉米种子萌发相关性状的遗传机制, 本研究对476份玉米自交系种子萌发相关的6个性状进行调查, 结合125万个(1.25M) SNP标记, 利用3种统计模型(Q, K, Q+K)进行全基因关联分析(GWAS)。结果表明K模型能够较好地评价吸胀前重量、吸胀前体积、吸胀后重量、吸胀后体积和吸胀体积5个性状; Q+K模型能更好地评价吸胀重量性状。基于这6个性状的最优模型的GWAS结果, 共检测到15个种子萌发相关性状的显著SNP, 15个SNP对应6个QTL, 集中分布在玉米第3、第6、第7和第10染色体上, QTL内单个SNP能解释的表型变异为5.09%~7.85%。其中5个QTL可在多个生物学重复中被检测到。以最显著SNP所在基因或附近基因作为QTL的候选基因, 共筛选到6个最可能的候选基因。GRMZM2G148411是吸胀后重量、吸胀重量和吸胀体积3个性状共同鉴定到的QTL候选基因, 根据基因的功能注释, 该基因编码一个包含TLD-domain的钙离子结合蛋白, 可能是一种调控种子休眠与萌发的信号分子。本研究鉴定的QTL为解析玉米种子萌发的遗传机制和相应功能标记的开发奠定了基础。
| [1] |
Ellis R H.Seed and seedling vigor in relation to crop growth and yield.Plant Growth Regul, 1992, 11: 249-255
doi: 10.1007/BF00024563 |
| [2] |
Candela H, Hake S.The art and design of genetic screens: maize.Nat Rev Genet, 2008, 9: 192-203
doi: 10.1038/nrg2291 pmid: 18250623 |
| [3] |
Below F E, Seebauer J R, Uribelarrea M, Schneerman M C, Moose S P.Physiological changes accompanying long-term selection for grain protein in maize. In: Janick J ed. Plant Breeding Reviews. New York: John Wiley & Sons, 2004. pp 133-151
doi: 10.1002/9780470650240.ch7 |
| [4] |
Klein R J, Zeiss C, Chew E Y, Tsai J Y, Sackler R S, Haynes C, Henning A K, SanGiovanni J P, Mane S M, Mayne S T, Bracken M B, Ferris F L, Ott J, Barnstable C, Hoh J. Complement factor H polymorphism in age-related macular degeneration. Science, 2005, 308: 385-389
doi: 10.1126/science.1109557 pmid: 15761122 |
| [5] | Atwell S, Huang Y S, Vilhjálmsson B J, Willems G, Horton M, Li Y, Meng D, Platt A, Tarone A M, Hu T T, Jiang R, Muliyati N W, Zhang X, Amer M A, Baxter I, Brachi B, Chory J, Dean C, Debieu M, de Meaux J, Ecker J R, Faure N, Kniskern J M, Jones J D, Michael T, Nemri A, Roux F, Salt D E, Tang C, Todesco M, Traw M B, Weigel D, Marjoram P, Borevitz J O, Bergelson J, Nordborg M. Genome-wide association study of 107 phenotypes in a common set of Arabidopsis thaliana inbred lines. Nature, 2010, 465: 627-631 |
| [6] |
Huang X H, Zhao Y, Wei X H, Li C Y, Wang A H, Zhao Q, Li W J, Guo Y L, Deng L W, Zhu C R, Fan D L, Lu Y Q, Weng Q J, Liu K Y, Zhou T Y, Jing Y F, Si L Z, Dong G J, Huang T, Lu T T, Feng Q, Qian Q, Li J Y, Han B.Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat Genet, 2011, 44: 32-39
doi: 10.1038/ng.1018 pmid: 22138690 |
| [7] |
Xiao Y J, Liu H J, Wu L J, Warburton M, Yan J B.Genome-wide association studies in maize: praise and stargaze.Mol Plant, 2017, 10: 359-374
doi: 10.1016/j.molp.2016.12.008 pmid: 28039028 |
| [8] |
Li H, Peng Z Y, Yang X H, Wang W D, Fu J J, Wang J H, Han Y J, Chai Y C, Guo T T, Yang N, Liu J, Warburton M L, Cheng Y B, Hao X M, Zhang P, Zhao J Y, Liu Y J, Wang G Y, Li J S, Yan J B.Genome-wide association study dissects the genetic architecture of oil biosynthesis in maize kernels. Nat Genet, 2013, 45: 43-50
doi: 10.1038/ng.2484 pmid: 23242369 |
| [9] |
Liu H J, Luo X, Niu L Y, Xiao Y J, Chen L, Liu J, Wang X G, Jin M L, Li W Q, Zhang Q H, Yan J B.Distant eQTLs and non-coding sequences play critical roles in regulating gene expression and quantitative trait variation in maize.Mol Plant, 2017, 10: 414-426
doi: 10.1016/j.molp.2016.06.016 pmid: 27381443 |
| [10] |
Huang J, Zhang J H, Li W Z, Hu W, Duan L C, Feng Y Z, Qiu F Z, Yue B.Genome-wide association analysis of ten chilling tolerance indices at the germination and seedling stages in maize.J Integr Plant Biol, 2013, 55: 735-744
doi: 10.1111/jipb.v55.8 |
| [11] |
Shi Y Y, Gao L L, Wu Z C, Zhang X J, Wang M M, Zhang C S, Zhang F, Zhou Y L, Li Z K.Genome-wide association study of salt tolerance at the seed germination stage in rice.BMC Plant Biol, 2017, 17: 92
doi: 10.1186/s12870-017-1044-0 pmid: 5450148 |
| [12] |
Kan G Z, Zhang W, Yang W M, Ma D Y, Zhang D, Hao D R, Hu Z B, Yu D Y.Association mapping of soybean seed germination under salt stress.Mol Genet Genomics, 2015, 290: 2147-2162
doi: 10.1007/s00438-015-1066-y pmid: 26001372 |
| [13] |
Finch-Savage W E, Leubner-Metzger G. Seed dormancy and the control of germination.New Phytol, 2006, 171: 501-523
doi: 10.1111/nph.2006.171.issue-3 |
| [14] |
Fu Z Y, Jin X N, Ding D, Li Y L, Fu Z J, Tang J H.Proteomic analysis of heterosis during maize seed germination.Proteomics, 2011, 11: 1462-1472
doi: 10.1002/pmic.201000481 pmid: 21365753 |
| [15] |
Hu S D, Lübberstedt T, Zhao G W, Lee M.QTL mapping of low-temperature germination ability in the maize IBM Syn4 RIL population.PLoS One, 2016, 11: e0152795
doi: 10.1371/journal.pone.0152795 pmid: 4816396 |
| [16] |
Han Z P, Ku L X, Zhang Z Z, Zhang J, Guo S L, Liu H V, Zhao R F, Ren Z Z, Zhang L K, Su H H, Dong L, Chen Y H.QTLs for seed vigor-related traits identified in maize seeds germinated under artificial aging conditions.PLoS One, 2014, 9: e92535
doi: 10.1371/journal.pone.0092535 pmid: 3961396 |
| [17] |
Yazdanpanah F, Hanson J, Hilhorst H, Bentsink L.Differentially expressed genes during the imbibition of dormant and after- ripened seeds—a reverse genetics approach.BMC Plant Biol, 2017, 17: 151-162
doi: 10.1186/s12870-017-1098-z pmid: 28893189 |
| [18] |
Li Y S, Yuan F, Wen Z H, Li Y H, Wang F, Zhu T, Zhuo W Q, Jin X, Wang Y D, Zhao H P, Pei Z M, Han S C.Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol, 2015, 15: 261-273
doi: 10.1186/s12870-015-0653-8 pmid: 4624379 |
| [19] |
Noblet A, Leymarie J, Bailly C.Chilling temperature remodels phospholipidome of Zea mays seeds during imbibition. Sci Rep, 2017, 7: 8886
doi: 10.1038/s41598-017-08904-z pmid: 28827663 |
| [20] |
Deng M, Li D Q, Luo J Y, Xiao Y J, Liu H J, Pan Q C, Zhang X H, Jin M L, Zhao M C, Yan J B.The genetic architecture of amino acids dissection by association and linkage analysis in maize.Plant Biotechnol J, 2017, 15: 1250-1263
doi: 10.1111/pbi.12712 pmid: 5595712 |
| [21] |
Wang M, Yan J B, Zhao J R, Song W, Zhang X B, Xiao Y N, Zheng Y L.Genome-wide association study (GWAS) of resistance to head smut in maize.Plant Sci, 2012, 196: 125-131
doi: 10.1016/j.plantsci.2012.08.004 pmid: 23017907 |
| [22] | Batak I, Devic M, Giba Z, Grubisic D, Poff K L, Konjevic R.The effects of potassium nitrate and NO-donors on phytochrome A- and phytochrome B-specific induced germination of Arabidopsis thaliana seeds. Seed Sci Res, 2002, 12: 253-259 |
| [23] | Bethke P C, Gubler F, Jacobsen J V, Jones R L.Dormancy of Arabidopsis seeds and barley grains can be broken by nitric oxide.Planta, 2004, 219: 847-855 |
| [24] |
Bethke P C, Libourel I G, Reinohl V, Jones R L.Sodium nitroprusside, cyanide, nitrite and nitrate break Arabidopsis seed dormancy in a nitric oxide-dependent manner.Planta, 2006, 223: 805-812
doi: 10.1007/s00425-005-0116-9 pmid: 16151848 |
| [25] |
Sarath G, Bethke P C, Jones R, Baird L M, Hou G, Mitchell R B.Nitric oxide accelerates seed germination in warm-season grasses.Planta, 2006, 223: 1154-1164
doi: 10.1007/s00425-005-0162-3 pmid: 16369800 |
| [26] | Fontaine O, Huault C, Pavis N, Billard J P.Dormancy breakage of Hordeum vulgare seeds: effects of hydrogen peroxide and scarification on glutathione level and glutathione reductase activity.Plant Physiol Biochem, 1994, 32: 677-683 |
| [27] |
El-Maarouf-Bouteau H, Bailly C. Oxidative signaling in seed germination and dormancy.Plant Signal Behav, 2008, 3: 175-182
doi: 10.4161/psb.3.3.5539 pmid: 2634111 |
| [28] |
Ishibashi Y, Kasa S, Sakamoto M, Aoki N, Kai K, Yuasa T, Hanada A, Yamaguchi S, Iwaya-Inoue M.A role for reactive oxygen species produced by NADPH oxidases in the embryo and aleurone cells in barley seed germination.PLoS One, 2015, 10: e0143173
doi: 10.1371/journal.pone.0143173 pmid: 26579718 |
| [29] |
Baek D, Cha J Y, Kang S, Park B, Lee H J, Hong H, Chun H J, Kim D H, Kim M C, Lee S Y, Yun D J.The Arabidopsisa zinc finger domain protein ARS1 is essential for seed germination and ROS homeostasis in response to ABA and oxidative stress.Front Plant Sci, 2015, 6: 963
doi: 10.3389/fpls.2015.00963 pmid: 4631831 |
| [30] |
Joseph M P, Papdi C, Kozma-Bognár L, Nagy I, López-Carbonell M, Rigó G, Koncz C, Szabados L.The Arabidopsis ZINC FINGER PROTEIN3 interferes with abscisic acid and light signaling in seed germination and plant development.Plant Physiol, 2014, 165: 1203-1220
doi: 10.1104/pp.113.234294 pmid: 24808098 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 陈玲玲, 李战, 刘亭萱, 谷勇哲, 宋健, 王俊, 邱丽娟. 基于783份大豆种质资源的叶柄夹角全基因组关联分析[J]. 作物学报, 2022, 48(6): 1333-1345. |
| [4] | 田甜, 陈丽娟, 何华勤. 基于Meta-QTL和RNA-seq的整合分析挖掘水稻抗稻瘟病候选基因[J]. 作物学报, 2022, 48(6): 1372-1388. |
| [5] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [6] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [7] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [8] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [9] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [10] | 孙思敏, 韩贝, 陈林, 孙伟男, 张献龙, 杨细燕. 棉花苗期根系分型及根系性状的关联分析[J]. 作物学报, 2022, 48(5): 1081-1090. |
| [11] | 于春淼, 张勇, 王好让, 杨兴勇, 董全中, 薛红, 张明明, 李微微, 王磊, 胡凯凤, 谷勇哲, 邱丽娟. 栽培大豆×半野生大豆高密度遗传图谱构建及株高QTL定位[J]. 作物学报, 2022, 48(5): 1091-1102. |
| [12] | 雷新慧, 万晨茜, 陶金才, 冷佳俊, 吴怡欣, 王家乐, 王鹏科, 杨清华, 冯佰利, 高金锋. 褪黑素与2,4-表油菜素内酯浸种对盐胁迫下荞麦发芽与幼苗生长的促进效应[J]. 作物学报, 2022, 48(5): 1210-1221. |
| [13] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [14] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [15] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
|
||