欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (3): 385-394.doi: 10.3724/SP.J.1006.2020.93021
马娟,曹言勇,王利锋,李晶晶,王浩,范艳萍,李会勇( )
)
Juan MA,Yan-Yong CAO,Li-Feng WANG,Jing-Jing LI,Hao WANG,Yan-Ping FAN,Hui-Yong LI()
摘要:
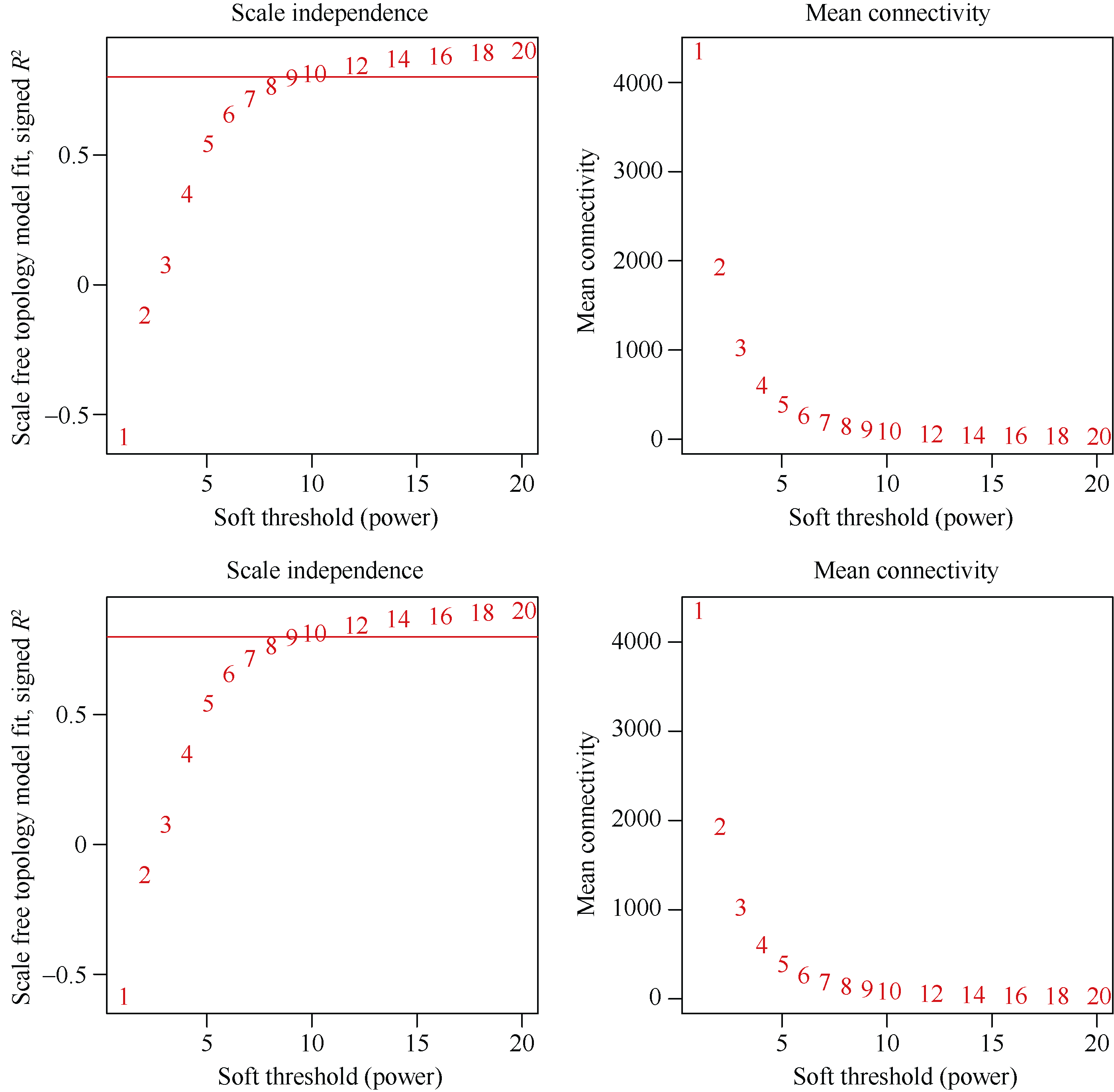
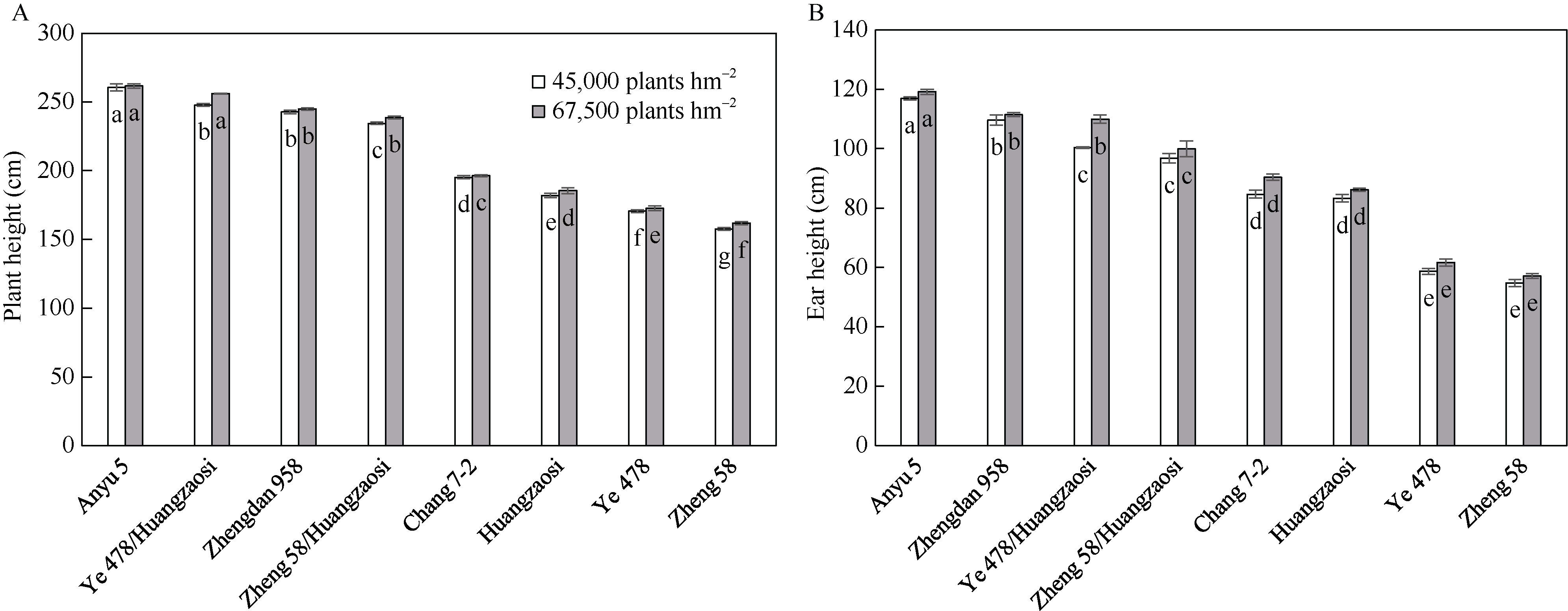
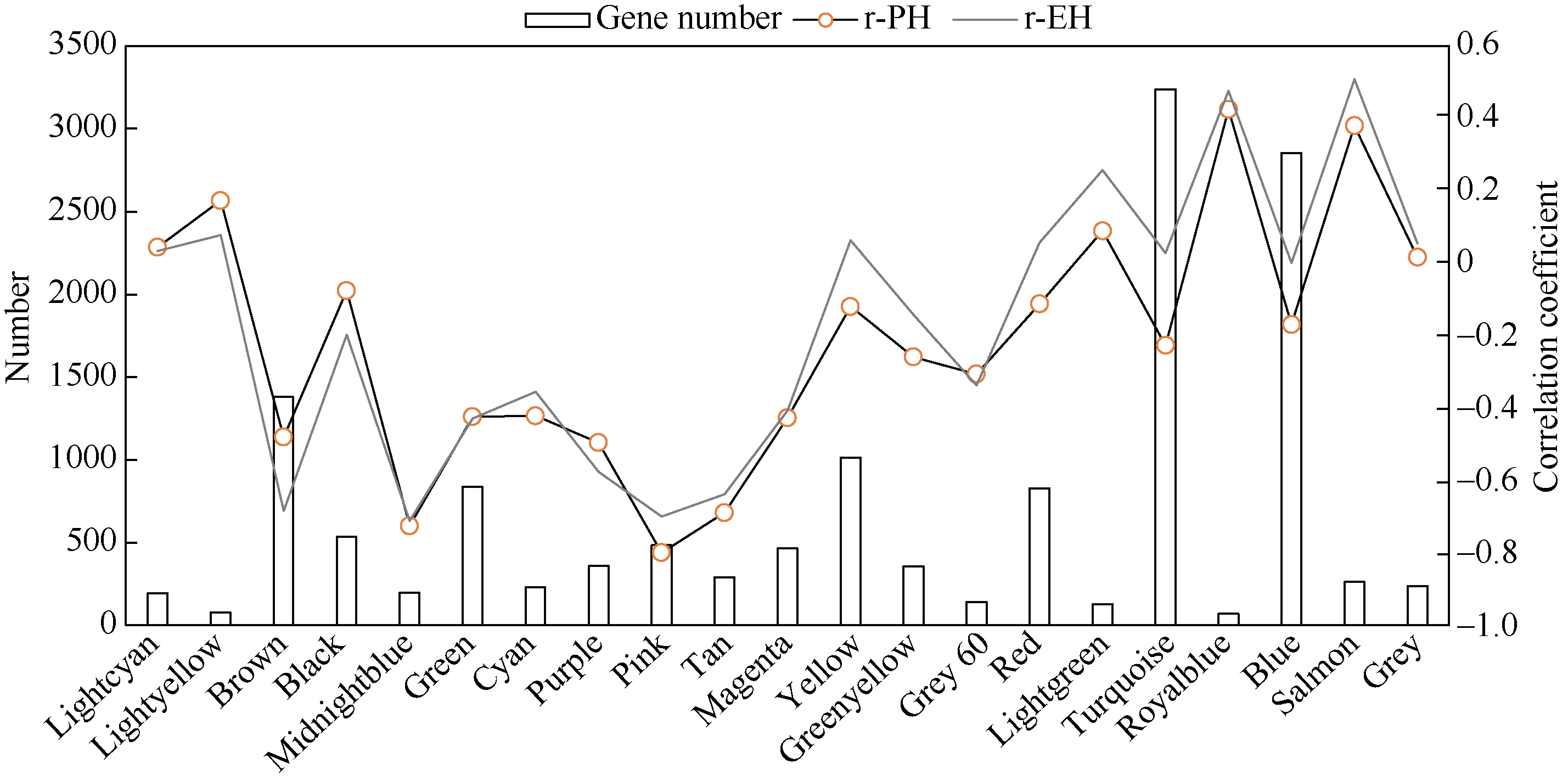
株高和穗位高是玉米株型的重要影响因子, 与产量性状紧密相关。加权基因共表达网络分析(weighted gene co-expression network analysis, WGCNA)是探索基因网络与特定性状间关联关系的重要方法, 为株高和穗位高相关基因的挖掘提供新途径。本研究利用郑58、掖478、昌7-2和黄早四及其组配的杂交种郑单958、安玉5号、郑58/黄早四和掖478/黄早四, 结合其在45,000株 hm -2和67,500株 hm -2条件下的转录组数据, 采用WGCNA构建了2种密度条件下的共表达网络, 分别得到24个和21个共表达模块, 并鉴定到与株高和穗位高显著且高度相关(相关系数的绝对值>0.50)的共表达模块15个, 其中两性状相同的模块共6个。基因功能富集分析结果表明, 株高和穗位高共表达模块主要参与生长发育、光合作用、响应光刺激、植物激素、碳水化合物合成/代谢等重要活动。根据模块内基因的连接度, 发现乙烯响应因子EREB14、硫胺素酶TENA2、磷酸甘油酸激酶PGK、谷胱甘肽转移酶GST2和琥珀酸脱氢酶SUDH7等是模块内的核心基因。通过构建其局部网络, 发现EREB14与已报道株高基因D8、DWF1和ZmGRF10以及C3H转录因子C3H35、C2C2-GATA转录因子GATA4和乙烯受体同源子ETR40等存在关联关系。此外, 已报道株高基因An1和GA20ox3也存在于共表达模块中。以这5个已报道株高基因为核心, 构建其基因网路, 发现生长素转录因子ARFTF7、ARFTF26、GST39、光合系统II氧进化多肽PspB2和光合系统I N亚基PasN1等与其存在关联。15个共表达模块和核心基因的挖掘以及基因生物学功能和互作网络的解析有助于揭示玉米株高和穗位高的遗传基础。
| [1] | 李清超, 李永祥, 杨钊钊, 刘成, 刘志斋, 李春辉, 彭勃, 张岩, 王迪, 谭巍巍, 孙宝成, 石云素, 宋燕春, 张志明, 潘光堂, 黎裕, 王天宇 . 基于多重相关RIL群体的玉米株高和穗位高QTL定位. 作物学报, 2013,39:1521-1529. |
| Li Q C, Li Y X, Yang Z Z, Liu C, Liu Z Z, Li C H, Peng B, Zhang Y, Wang D, Tan W W, Sun B C, Shi S Y, Song C Y, Zhang Z M, Pan G T, Li Y, Wang T Y . QTL mapping for plant height and ear height by using multiple related RIL populations in maize. Acta Agron Sin, 2013,39:1521-1529 (in Chinese with English abstract). | |
| [2] | 何坤辉, 常立国, 崔婷婷, 渠建洲, 郭东伟, 徐淑兔, 张兴华, 张仁和, 薛吉全, 刘建超 . 多环境下玉米株高和穗位高的QTL定位. 中国农业科学, 2016,49:1443-1452. |
| He K H, Chang L G, Cui T T, Qu J Z, Guo D W, Xu S T, Zhang X H, Zhang R H, Xue J Q, Liu J C . Mapping QTL for plant height and ear height in maize under multi-environments. Sci Agric Sin, 2016,49:1443-1452 (in Chinese with English abstract). | |
| [3] | 刘坤, 张雪海, 孙高阳, 闫鹏帅, 郭海平, 陈思远, 薛亚东, 郭战勇, 谢惠玲, 汤继华, 李卫华 . 玉米株型相关性状的全基因组关联分析. 中国农业科学, 2018,51:821-834. |
| Liu K, Zhang X H, Sun G Y, Yan P S, Guo H P, Chen S Y, Xue Y D, Guo Z Y, Xie H L, Tang J H, Li W H . Genome-wide association studies of plant type traits in maize. Sci Agric Sin, 2018,51:821-834 (in Chinese with English abstract). | |
| [4] | 李凯, 张晓祥, 管中荣, 沈亚欧, 潘光堂 . 玉米株高和穗位高的全基因组关联分析. 玉米科学, 2017,25(6):1-7. |
| Li K, Zhang X X, Guan Z R, Shen Y O, Pan G T . Genome-wide association analysis of plant height and ear height in maize. J Maize Sci, 2017,25(6):1-7 (in Chinese with English abstract). | |
| [5] | Li X, Zhou Z, Ding J, Wu Y, Zhou B, Wang R, Ma J, Wang S, Zhang X, Xia Z, Chen J, Wu J . Combined linkage and association mapping reveals QTL and candidate genes for plant and ear height in maize. Front Plant Sci, 2016,7:833. |
| [6] | Weng J, Xie C, Hao Z, Wang J, Liu C, Li M, Zhang D, Bai L, Zhang S, Li X . Genome-wide association study identifies candidate genes that affect plant height in Chinese elite maize (Zea mays L.) inbred lines. PLoS One, 2011,6:e29229. |
| [7] | Fujioka S, Yamane H, Spray C R, Gaskin P, Macmillan J, Phinney B O, Takahashi N . Qualitative and quantitative analyses of gibberellins in vegetative shoots of normal, dwarf-1, dwarf-2, dwarf-3, and dwarf-5 seedlings of Zea mays L. Plant Physiol, 1988,88:1367-1372. |
| [8] | Winkler R G, Helentjaris T . The maize Dwarf3 gene encodes acytochrome P450-mediated early step in gibberellin biosynthesis. Plant Cell, 1995,7:1307-1317. |
| [9] | Thornsberry J M, Goodman M M, Doebley J, Kresovich S, Nielsen D, Buckler E S . Dwarf8 polymorphisms associate with variation in flowering time. Nat Genet, 2001,28:286-289. |
| [10] | Lawit S J, Wych H M, Xu D, Kundu S, Tomes D T . Maize DELLA proteins dwarf plant8 and dwarf plant9 as modulators of plant development. Plant Cell Physiol, 2010,51:1854-1868. |
| [11] | Multani D S, Briggs S P, Chamberlin M A, Blakeslee J J, Murphy A S, Johal G S . Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants. Science, 2003,302:81-84. |
| [12] | Zhang B, Horvath S. A general framework for weighted gene co-expression network analysis. Stat Appl Genet Mol Biol, 2005, 4: Article 17. |
| [13] | Zhang X, Hirsch C N, Sekhon R S, De Leon N, Kaeppler S M . Evidence for maternal control of seed size in maize from phenotypic and transcriptional analysis. J Exp Bot, 2016,67:1907-1917. |
| [14] | Ma J, Zhang D, Cao Y, Wang L, Li J, Lübberstedt T, Wang T, Li Y, Li H . Heterosis-related genes under different planting densities in maize (Zea mays L.). J Exp Bot, 2018,69:5077-5087. |
| [15] | Zhan J, Thakare D, Ma C, Lloyd A, Nixon N M, Arakaki A M, Burnett W J, Logan K O, Wang D, Wang X, Drews G N, Yadegari R . RNA sequencing of laser-capture microdissected compartments of the maize kernel identifies regulatory modules associated with endosperm cell differentiation. Plant Cell, 2015,27:513-531. |
| [16] | 杨宇昕, 桑志勤, 许诚, 代文双, 邹枨 . 利用WGCNA进行玉米花期基因共表达模块鉴定. 作物学报, 2019,45:161-174. |
| Yang Y X, Sang Z Q, Xu C, Dai W S, Zou C . Identification of maize flowering gene co-expression modules by WGCNA. Acta Agron Sin, 2019,45:161-174 (in Chinese with English abstract). | |
| [17] | Peng H, He X, Gao J, Ma H, Zhang Z, Shen Y, Pan G, Lin H . Transcriptomic changes during maize roots development responsive to Cadmium (Cd) pollution using comparative RNA seq- based approach. Biochem Biophys Res Commun, 2015,464:1040-1047. |
| [18] | Thirunavukkarasu N, Hossain F, Mohan S, Shiriga K, Mittal S, Sharma R, Singh R K, Gupta H S . Genome-wide expression of transcriptomes and their co-expression pattern in subtropical maize (Zea mays L.) under waterlogging stress. PLoS One, 2013,8:e70433. |
| [19] | Lyu Y, Liang Z, Ge M, Qi W, Zhang T, Lin F, Peng Z, Zhao H . Genome-wide identification and functional prediction of nitrogen-responsive intergenic and intronic long non-coding RNAs in maize ( Zea mays L.). BMC Genomics, 2016,17:350. |
| [20] | Zhang S, Yang W, Zhao Q, Zhou X, Jiang L, Ma S, Liu X, Li Ye, Zhang C, Fan Y, Chen R . Analysis of weighted co-regulatory networks in maize provides insights into new genes and regulatory mechanisms related to inositol phosphate metabolism. BMC Genomics, 2016,17:129-146. |
| [21] | Tao Y, Zheng J, Xu Z, Zhang X, Zhang K, Wang G . Functional analysis of ZmDWF1, a maize homolog of the Arabidopsis brassinosteroids biosynthetic DWF1/DIM gene. Plant Sci, 2004,167:741-751. |
| [22] | Wu L, Zhang D, Xue M, Qian J, He Y, Wang S . Overexpression of the maize GRF10, an endogenous truncated growth regulating factor protein, leads to reduction in leaf size and plant height. J Integr Plant Biol, 2014,56:1053-1063. |
| [23] | Hartwig T, Chuck G S, Fujioka S, Klempien A, Weizbauer R, Potluri D P, Choe S, Johal G S, Schulz B . Brassinosteroid control of sex determination in maize. Proc Natl Acad Sci USA, 2011,108:19814-19819. |
| [24] | Tamotsu H, Rod W K, Chris A H, Masaji K . The involvement of gibberellin 20-oxidase genes in phytochrome-regulated petiole elongation of Arabidopsis. Plant Physiol, 2005,138:1106-1116. |
| [25] | Zhao W, Langfelder P, Fuller T, Dong J, Li A, Hovarth S . Weighted gene coexpression network analysis: state of the art. J Biopharm Stat, 2010,20:281-300. |
| [26] | Wang H, Gu L, Zhang X, Liu M, Jiang H, Cai R, Zhao Y, Cheng B . Global transcriptome and weighted gene co-expression network analyses reveal hybrid-specific modules and candidate genes related to plant height development in maize. Plant Mol Biol, 2018,98:187-203. |
| [27] | Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K . AP2/ERF family transcription factors in plant abiotic stress responses. BBA-Gene Regul Mech, 2012,1819:86-96. |
| [28] | Hinz M, Wilson I W, Yang J, Buerstenbinder K, Llewellyn D, Dennis E S, Sauter M, Dolferus R . Arabidopsis RAP2: 2. An ethylene response transcription factor that is important for hypoxia survival. Plant Physiol, 2010,153:757-772. |
| [29] | Licausi F, Ohme Takagi M, Perata P . APETALA2/ethylene responsive factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs. New Phytol, 2013,199:639-649. |
| [30] | Cassani E, Bertolini E, Cerino B F, Landoni M, Gavina D, Sirizzotti A, Pilu R . Characterization of the first dominant dwarf maize mutant carrying a single amino acid insertion in the VHYNP domain of the dwarf8 gene. Mol Breed, 2009,24:375-385. |
| [31] | Teng F, Zhai L, Liu R, Bai W, Wang L, Huo D, Tao Y, Zheng Y, Zhang Z . ZmGA3ox2, a candidate gene for a major QTL, qPH3.1, for plant height in maize. Plant J, 2013,73:405-416. |
| [32] | 郑雷, 周羽, 曾兴, 邸宏, 翁建峰, 李新海, 王振华 . 玉米株高QTL定位研究进展. 作物杂志, 2016, ( 2):8-13. |
| Zheng L, Zhou Y, Zeng X, Di H, Weng J F, Li X H, Wang Z H . QTL Mapping of plant height in maize. Crops, 2016, ( 2):8-13 (in Chinese with English abstract). |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [4] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [5] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [6] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [7] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [8] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [9] | 于春淼, 张勇, 王好让, 杨兴勇, 董全中, 薛红, 张明明, 李微微, 王磊, 胡凯凤, 谷勇哲, 邱丽娟. 栽培大豆×半野生大豆高密度遗传图谱构建及株高QTL定位[J]. 作物学报, 2022, 48(5): 1091-1102. |
| [10] | 王泽, 周钦阳, 刘聪, 穆悦, 郭威, 丁艳锋, 二宫正士. 基于无人机和地面图像的田间水稻冠层参数估测与评价[J]. 作物学报, 2022, 48(5): 1248-1261. |
| [11] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [12] | 孔垂豹, 庞孜钦, 张才芳, 刘强, 胡朝华, 肖以杰, 袁照年. 不同施肥水平下丛枝菌根真菌对甘蔗生长及养分相关基因共表达网络的影响[J]. 作物学报, 2022, 48(4): 860-872. |
| [13] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [14] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [15] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
|
||