欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (8): 1437-1449.doi: 10.3724/SP.J.1006.2021.01077
王艳朋( ), 凌磊, 张文睿, 王丹, 郭长虹*()
), 凌磊, 张文睿, 王丹, 郭长虹*()
WANG Yan-Peng(), LING Lei, ZHANG Wen-Rui, WANG Dan, GUO Chang-Hong*()
摘要:
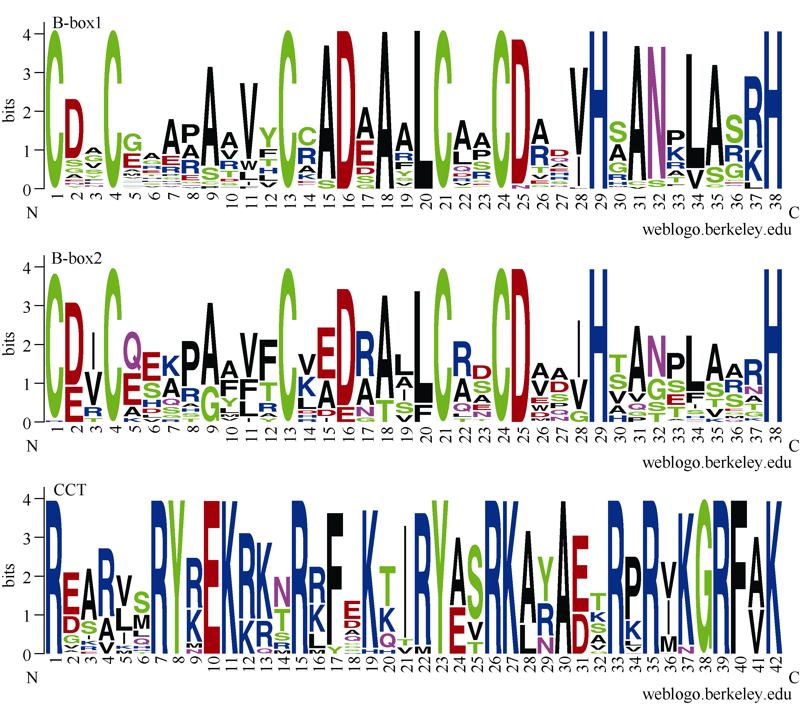
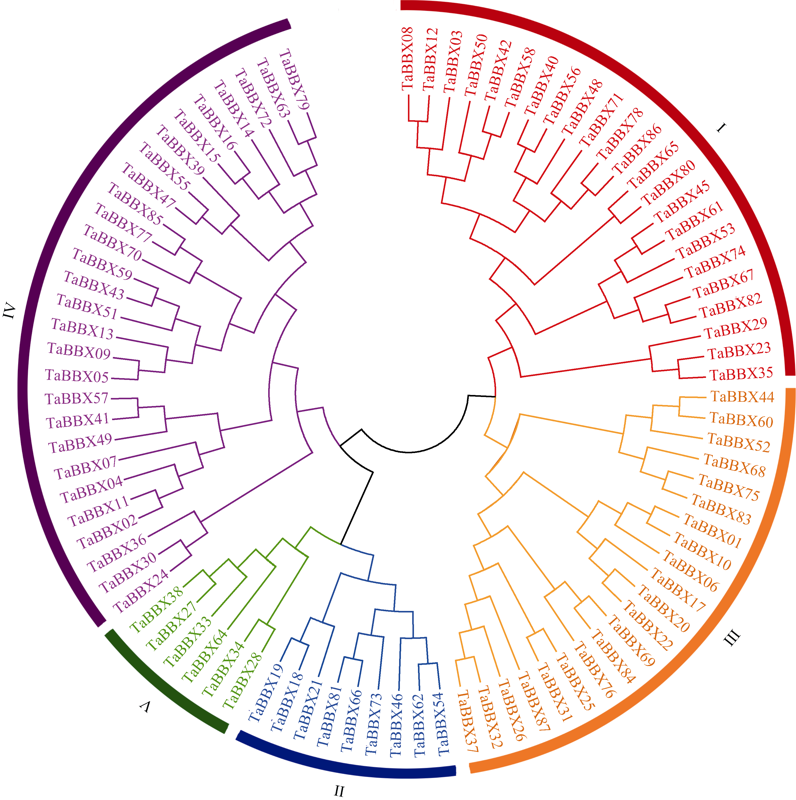
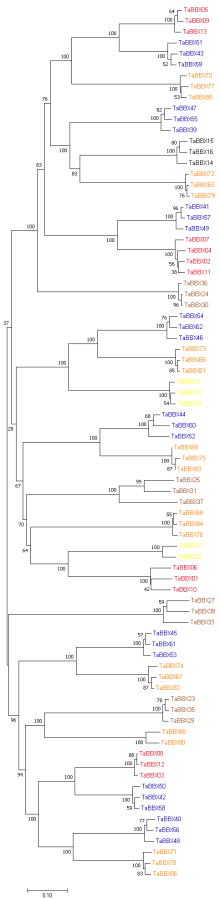
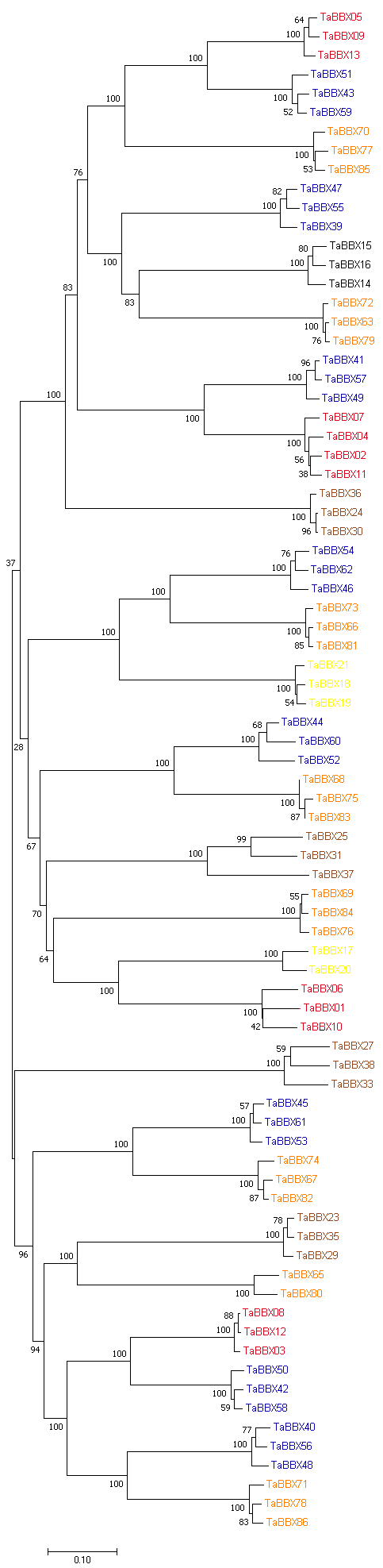
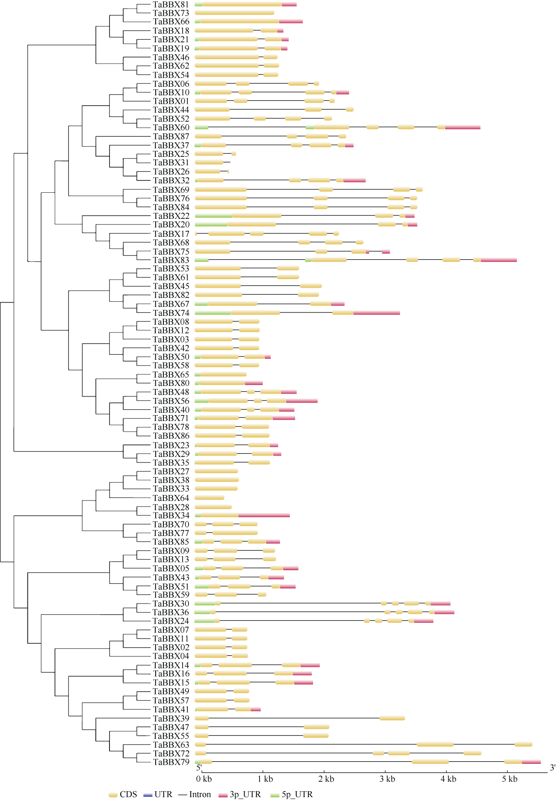
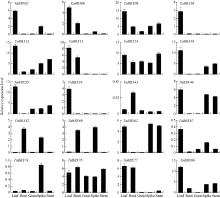
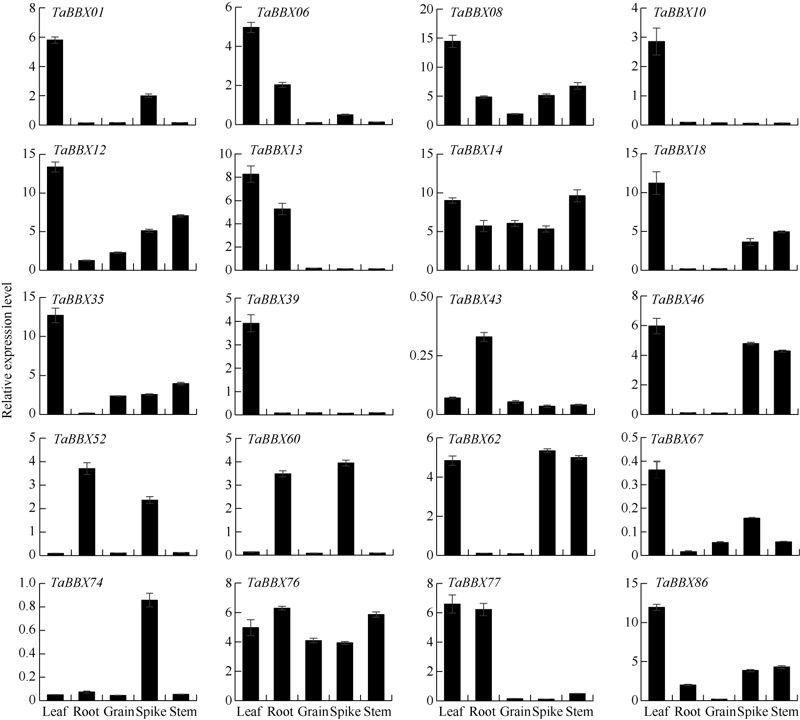
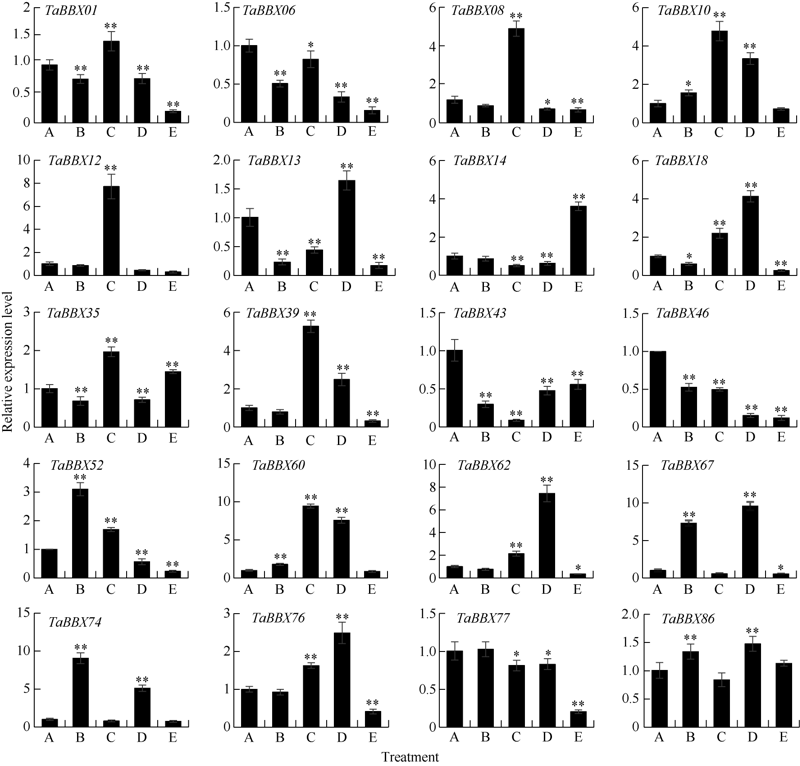
B-box (BBX)是一类含有1个或2个B-box结构域的锌指蛋白, 在植物生长发育中起着重要作用。本研究明确小麦B-box转录因子的数量、基因结构和分类进化关系, 研究各基因成员在不同组织中的特异性表达以及对非生物胁迫的响应。从小麦全基因组中鉴定得到87个B-box基因家族成员, 所有TaBBXs蛋白均含有B-box结构域。TaBBXs编码146~489个氨基酸, 理论等电点为4.32~10.42。染色体定位分析表明, TaBBXs分布在除1A、1B和1D之外的18条小麦染色体上。通过系统发育分析将TaBBXs划分为5个亚家族, 有0~4个内含子。在同组内同一个系统进化树分支中的亚族成员具有高度相似的基因结构。qRT-PCR分析的20个TaBBXs基因, 具有不同的组织表达模式, 16个基因在叶中有较高表达, TaBBX10和TaBBX39仅在叶中有较高表达, 而TaBBX74在穗中表达, TaBBX43在根中特异性表达。在不同逆境胁迫下, TaBBXs呈现不同表达模式, 11个基因在低温胁迫后上调表达, 12个基因在ABA处理后下调表达, 盐胁迫后10个基因出现上调表达, 干旱胁迫后7个基因出现下调表达, TaBBX10、TaBBX39、TaBBX60、TaBBX67和TaBBX74基因在2种或2种以上胁迫下有显著的上调表达。
| [1] | 解松峰, 吉万全, 张耀元, 张俊杰, 胡卫国, 李俊, 王长有, 张宏, 陈春环. 小麦重要产量性状的主基因+多基因混合遗传分析. 作物学报, 2020,46:365-384. |
| Xie S F, Ji W Q, Zhang Y Y, Zhang J J, Hu W G, Li J, Wang C Y, Zhang H, Chen C H. Genetic effects of important yield traits analyzed by mixture model of major gene plus polygene in wheat. Acta Agron Sin, 2020,46:365-384 (in Chinese with English abstract). | |
| [2] | 刘登才, 张连全, 郝明, 黄林, 甯顺腙, 袁中伟, 姜博, 颜泽洪, 伍碧华, 郑有良. 小麦族的基因组显性及其育种学意义. 作物学报, 2020,46:1465-1473. |
| Liu D C, Zhang L Q, Hao M, Huang L, Ning S Z, Yuan Z W, Jiang B, Yan Z H, Wu B H, Zheng Y L. Genome dominance and the breeding significance in Triticeae. Acta Agron Sin, 2020,46:1465-1473 (in Chinese with English abstract). | |
| [3] |
Choulet F, Alberti A, Theil S, Glover N, Barbe V, Daron J, Pingault L, Sourdille P, Couloux A, Paux E, Leroy P, Mangenot S, Guilhot N, Le Gouis J, Balfourier F, Alaux m, Jamilloux V, Poulain J, Durand C, Bellec A, Gaspin C, Safar J, Dolezel J, Rogers J, Vandepoele K, Aury J M, Mayer K, Berges H, Quesneville H, Wincker P, Feuillet C. Structural and functional partitioning of bread wheat chromosome 3B. Science, 2014,345:1249721.
doi: 10.1126/science.1249721 |
| [4] |
IWGSC. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science, 2018,361:eaar7191.
doi: 10.1126/science.aar7191 |
| [5] | 茹京娜, 于太飞, 陈隽, 陈明, 周永斌, 马有志, 徐兆师, 闵东红. 小麦锌指转录因子TaDi19A对低温的响应及其互作蛋白的筛选. 中国农业科学, 2017,50:2411-2422. |
| Ru J N, Yu T F, Chen J, Chen M, Zhou Y B, Ma Y Z, Xu Z S, Min D H. Response of wheat Zinc-Finger transcription factor TaDi19A to cold and its screening of interacting proteins. Sci Agric Sin, 2017,50:2411-2422 (in Chinese with English abstract). | |
| [6] |
Klug A, Schwabe J W. Protein motifs 5. Zinc fingers. FASEB J, 1995,9:597-604.
pmid: 7768350 |
| [7] |
Putterill J, Robson F, Lee K, Simon R, Coupland G. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell, 1995,80:847.
pmid: 7697715 |
| [8] |
Chang C S, Maloof J N, Wu S H. COP1-mediated degradation of BBX22/LZF1 optimizes seedling development in Arabidopsis. Plant Physiol, 2011, 156: 228-239.
doi: 10.1104/pp.111.175042 |
| [9] |
Gonzalezschain N D, Diazmendoza M, Zurczak M, Suarezlopez P. Potato CONSTANS is involved in photoperiodic tuberization in a graft-transmissible manner. Plant J, 2012,70:678-690.
doi: 10.1111/tpj.2012.70.issue-4 |
| [10] |
Crocco C D, Holm M, Yanovsky M J, Botto J F. Function of B-box under shade. Plant Signal Behav, 2011,6:101-104.
doi: 10.4161/psb.6.1.14185 |
| [11] |
Wang Q, Tu X, Zhang J, Chen X, Rao L. Heat stress-induced BBX18 negatively regulates the thermotolerance in Arabidopsis. Mol Biol Rep, 2013,40:2679-2688.
doi: 10.1007/s11033-012-2354-9 |
| [12] |
Fan X Y, Sun Y, Cao D M, Bai M Y, Luo X M, Yang H J, Wei C Q, Zhu S W, Sun Y, Chong K. BZS1, a B-box protein, promotes photomorphogenesis downstream of both Brassinosteroid and light signaling pathways. Mol Plant, 2012,5:591-600.
doi: 10.1093/mp/sss041 |
| [13] |
Bai M J, Sun J J, Liu J Y, Ren H R, Wang K, Wang Y L, Wang C Q, Dehesh K. The B-box protein BBX19 suppresses seed germination via induction of ABI5. Plant J, 2019,99:1192-1202.
doi: 10.1111/tpj.v99.6 |
| [14] |
Yadav A, Lingwan M, Yadukrishnan P, Masakapalli S K, Datta S. BBX31 promotes hypocotyl growth, primary root elongation and UV-B tolerance in Arabidopsis. Plant Signal Behav, 2019,14:e1588672.
doi: 10.1080/15592324.2019.1588672 |
| [15] |
Gangappa S N, Crocco C D, Johansson H, Datta S, Hettiarachchi C, Holm M, Botto J F. The Arabidopsis B-box protein BBX25 interacts with HY5, negatively regulating BBX22 expression to suppress seedling photomorphogenesis. Plant Cell, 2013,25:1243-1257.
doi: 10.1105/tpc.113.109751 |
| [16] |
An J P, Wang X F, Espley R V, Lin W K, Bi S Q, You C X, Hao Y J. An apple B-box protein MdBBX37 modulates anthocyanin biosynthesis and hypocotyl elongation synergistically with MdMYBs and MdHY5. Plant Cell Physiol, 2020,61:130-143.
doi: 10.1093/pcp/pcz185 |
| [17] |
Lippuner V, Cyert M S, Gasser C S. Two classes of plant cDNA clones differentially complement yeast calcineurin mutants and increase salt tolerance of wild-type yeast. J Biol Chem, 1996,271:12859-12866.
pmid: 8662738 |
| [18] |
Nagaoka S, Takano T. Salt tolerance-related protein STO binds to a MYB transcription factor homologue and confers salt tolerance in Arabidopsis. J Exp Bot, 2003,54:2231-2237.
doi: 10.1093/jxb/erg241 |
| [19] |
Min J H, Chung J S, Lee K H, Kim C S. The CONSTANS-like 4 transcription factor, AtCOL4, positively regulates abiotic stress tolerance through an abscisic acid-dependent manner in Arabidopsis. J Integr Plant Biol, 2015,57:313-324.
doi: 10.1111/jipb.12246 |
| [20] |
Liu X, Li R, Dai Y Q, Yuan L, Sun Q H, Zhang S Z, Wang X Y. A B-box zinc finger protein, MdBBX10, enhanced salt and drought stresses tolerance in Arabidopsis. Plant Mol Biol, 2019,99:437-447.
doi: 10.1007/s11103-019-00828-8 |
| [21] |
Liu X, Dai Y Q, Li R, Yuan L, Chen X S, Wang X Y. Members of B-box protein family from Malus domestica enhanced abiotic stresses tolerance in Escherichia coli. Mol Biotechnol, 2019,61:421-426.
doi: 10.1007/s12033-019-00172-8 |
| [22] | 王日红, 宋敏燕, 王然, 杨英杰. 山梨B-box基因PuBBX24表达特性及其在童期调控中的功能分析. 园艺学报, 2019,46:1458-1472. |
| Wang R H, Song M Y, Wang R, Yang Y J. Expression characteristics of B-box gene PuBBX24 and its function in juvenile regulation. Acta Hortic Sin, 2019,46:1458-1472 (in Chinese with English abstract). | |
| [23] |
Khanna R, Wu S H. The Arabidopsis B-box zinc finger family. Plant Cell, 2009,21:3416.
doi: 10.1105/tpc.109.069088 |
| [24] |
Huang J Y, Zhao X B, Weng X Y, Wang L, Xie W B. The rice B-box zinc finger gene family: genomic identification, characterization, expression profiling and diurnal analysis. PLoS One, 2012,7:e48242.
doi: 10.1371/journal.pone.0048242 |
| [25] |
Gangappa S N, Botto J F. The BBX family of plant transcription factors. Trends Plant Sci, 2014,19:460-470.
doi: 10.1016/j.tplants.2014.01.010 |
| [26] | Chu Z, Wang X, Li Y, Yu H, Li J, Lu Y, Li H, Ouyang B. Genomic organization, phylogenetic and expression analysis of the B-box gene family in tomato. Front Plant Sci, 2016,7:1552. |
| [27] |
Talar U, Kiełbowicz-Matuk A, Czarnecka J, Rorat T. Genome- wide survey of B-box proteins in potato (Solanum tuberosum)—Identification, characterization and expression patterns during diurnal cycle, etiolation and de-etiolation. PLoS One, 2017,12:e0177471.
doi: 10.1371/journal.pone.0177471 |
| [28] |
Shalmani A, Fan S, Jia P, Li G, Muhammad I, Li Y, Sharif R, Dong F, Zuo X, Li K. Genome identification of B-box gene family members in seven rosacea species and their expression analysis in response to flower induction in Malus domestica. Molecules, 2018,23:1763.
doi: 10.3390/molecules23071763 |
| [29] |
Liu X, Li R, Dai Y Q, Chen X S, Wang X Y. Genome-wide identification and expression analysis of the B-box gene family in the apple (Malus domestica Borkh.) genome. Mol Genet Genome, 2017,293:1-13.
doi: 10.1007/s00438-017-1370-9 |
| [30] |
Cao Y, Han Y, Meng D, Li D, Jiao C, Jin Q, Lin Y, Cai Y. B-box genes: genome-wide identification, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri Rehd.). BMC Plant Biol, 2017,17:156.
doi: 10.1186/s12870-017-1105-4 |
| [31] |
Wei H R, Wang P P, Chen J Q, Li C J, Wang Y Z, Yuan Y B, Fang J G, Leng X P. Genome-wide identification and analysis of B-box gene family in grapevine reveal its potential functions in berry development. BMC Plant Biol, 2020,20:72.
doi: 10.1186/s12870-020-2239-3 |
| [32] |
Crocco C D, Botto J F. BBX proteins in green plants: insights into their evolution, structure, feature and functional diversification. Gene, 2013,531:44-52.
doi: 10.1016/j.gene.2013.08.037 |
| [33] |
Zou Z Y, Wang R H, Wang R, Yang S L, Yang Y J. Genome-wide identification, phylogenetic analysis, and expression profiling of the BBX family genes in pear. J Hortic Sci Biotechnol, 2017,93:37-50.
doi: 10.1080/14620316.2017.1338927 |
| [34] |
Magadum S, Banerjee U, Murugan P, Gangapur D, Ravikesavan R. Gene duplication as a major force in evolution. J Genet, 2013,92:155-161.
doi: 10.1007/s12041-013-0212-8 |
| [35] |
Cannon S B, Mitra A, Baumgarten A, Young N D, May G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol, 2004,4:10.
doi: 10.1186/1471-2229-4-10 |
| [36] |
Moore R C, Purugganan M D. The early stages of duplicate gene evolution. Proc Natl Acad Sci USA, 2003,100:15682-15687.
doi: 10.1073/pnas.2535513100 |
| [37] |
Abdullah S, Jing X Q, Shi Y, Izhar M, Zhou M R, Wei X Y, Chen Q Q, Li W Q, Liu W T, Chen K M. Characterization of B-box gene family and their expression profiles under hormonal, abiotic and metal stresses in Poaceae plant. BMC Genome, 2019,20:27.
doi: 10.1186/s12864-018-5336-z |
| [38] |
Hassidim M, Harir Y, Yakir E, Kron I, Green R M. Over- expression of CONSTANS-LIKE 5 can induce flowering in short-day grown Arabidopsis. Planta, 2009,230:481-491.
doi: 10.1007/s00425-009-0958-7 pmid: 19504268 |
| [39] |
Cheng X F, Wang Z Y. Overexpression of COL9, a CONSTANS-LIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J, 2005,43:758-768.
doi: 10.1111/tpj.2005.43.issue-5 |
| [40] | Li W, Wang J, Sun Q, Li W, Yu Y, Zhao M, Meng Z. Expression analysis of genes encoding double B-box zinc finger proteins in maize. Funct Integr Genome, 2017,17:653-666. |
| [41] |
Rengasamy P. World salinization with emphasis on Australia. J Exp Bot, 2006,57:1017-1023.
pmid: 16510516 |
| [42] |
Cramer G R, Urano K, Delrot S, Pezzotti M, Shinozaki K. Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biol, 2011,11:163.
doi: 10.1186/1471-2229-11-163 pmid: 22094046 |
| [43] |
Yang Y, Ma C, Xu Y, Wei Q, Imtiaz M, Lan H, Gao S, Cheng L, Wang M, Fei Z. A zinc finger protein regulates flowering time and abiotic stress tolerance in chrysanthemum by modulating gibberellin biosynthesis. Plant Cell, 2014,26:2038-2354.
doi: 10.1105/tpc.114.124867 |
| [44] |
刘焱, 邢立静, 李俊华, 戴绍军. 水稻含有B-box锌指结构域的OsBBX25蛋白参与植物对非生物胁迫的响应. 植物学报, 2012,47:366-378.
doi: 10.3724/SP.J.1259.2012.00366 |
| Liu Y, Xing L J, Li J H, Dai S J. Rice B-box zinc finger protein OsBBX25 is involved in the abiotic response. Chin Bull Bot, 2012,47:366-378 (in Chinese with English abstract). |
| [1] | 陈玲玲, 李战, 刘亭萱, 谷勇哲, 宋健, 王俊, 邱丽娟. 基于783份大豆种质资源的叶柄夹角全基因组关联分析[J]. 作物学报, 2022, 48(6): 1333-1345. |
| [2] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [3] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [4] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [5] | 孙思敏, 韩贝, 陈林, 孙伟男, 张献龙, 杨细燕. 棉花苗期根系分型及根系性状的关联分析[J]. 作物学报, 2022, 48(5): 1081-1090. |
| [6] | 姚晓华, 王越, 姚有华, 安立昆, 王燕, 吴昆仑. 青稞新基因HvMEL1 AGO的克隆和条纹病胁迫下的表达[J]. 作物学报, 2022, 48(5): 1181-1190. |
| [7] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [8] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
| [9] | 刘运景, 郑飞娜, 张秀, 初金鹏, 于海涛, 代兴龙, 贺明荣. 宽幅播种对强筋小麦籽粒产量、品质和氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 716-725. |
| [10] | 马红勃, 刘东涛, 冯国华, 王静, 朱雪成, 张会云, 刘静, 刘立伟, 易媛. 黄淮麦区Fhb1基因的育种应用[J]. 作物学报, 2022, 48(3): 747-758. |
| [11] | 徐龙龙, 殷文, 胡发龙, 范虹, 樊志龙, 赵财, 于爱忠, 柴强. 水氮减量对地膜玉米免耕轮作小麦主要光合生理参数的影响[J]. 作物学报, 2022, 48(2): 437-447. |
| [12] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [13] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [14] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
| [15] | 马博闻, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑, 姜东. 花前渍水锻炼调控花后小麦耐渍性的生理机制研究[J]. 作物学报, 2022, 48(1): 151-164. |
|
||