欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (1): 63-75.doi: 10.3724/SP.J.1006.2022.01100
孟颖1( ), 邢蕾蕾1, 曹晓红1, 郭光艳1, 柴建芳2,*(), 秘彩莉1,*()
), 邢蕾蕾1, 曹晓红1, 郭光艳1, 柴建芳2,*(), 秘彩莉1,*()
MENG Ying1(), XING Lei-Lei1, CAO Xiao-Hong1, GUO Guang-Yan1, CHAI Jian-Fang2,*(), BEI Cai-Li1,*()
摘要:
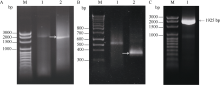
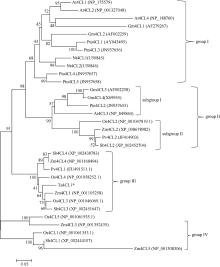
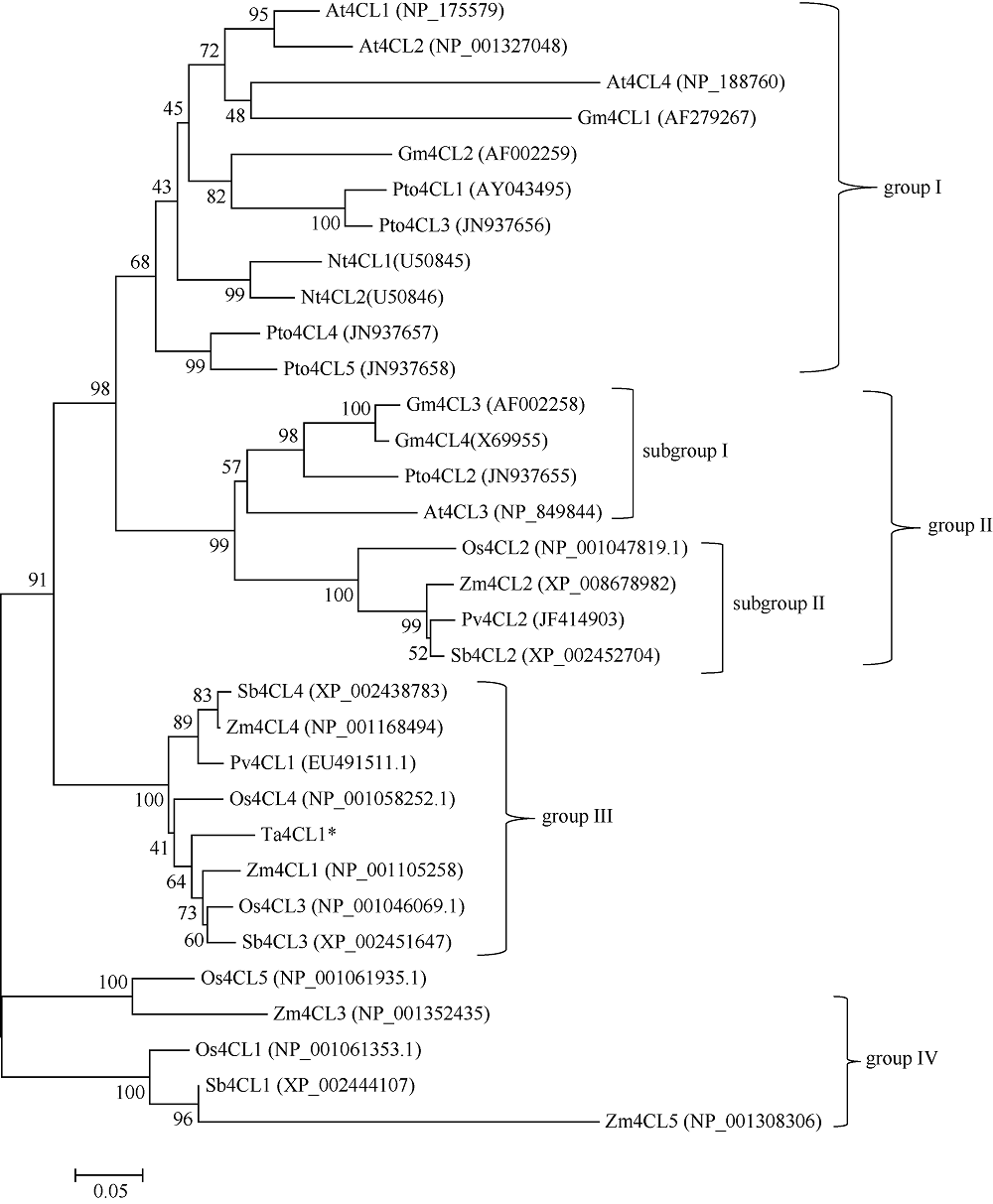
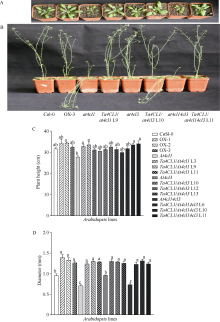
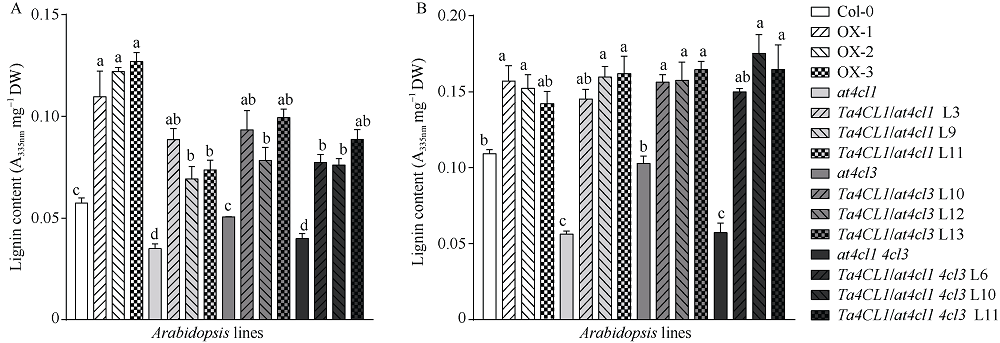
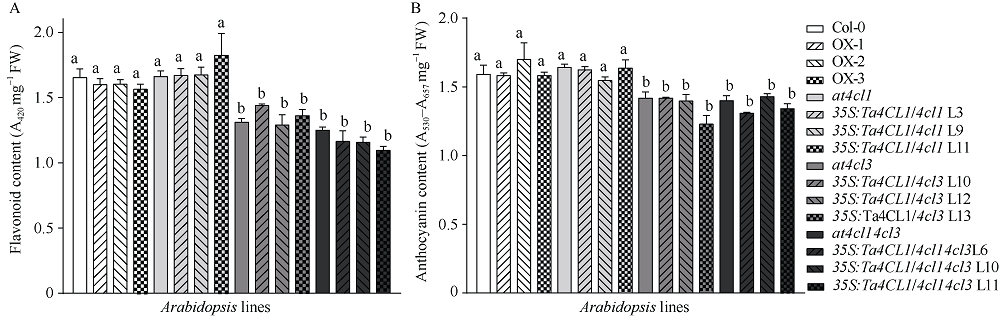
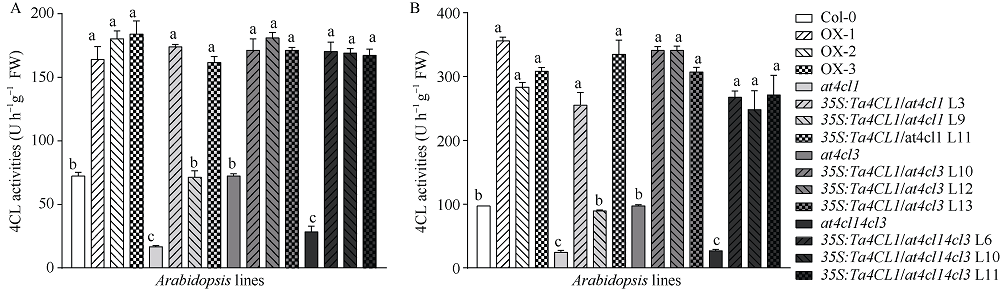
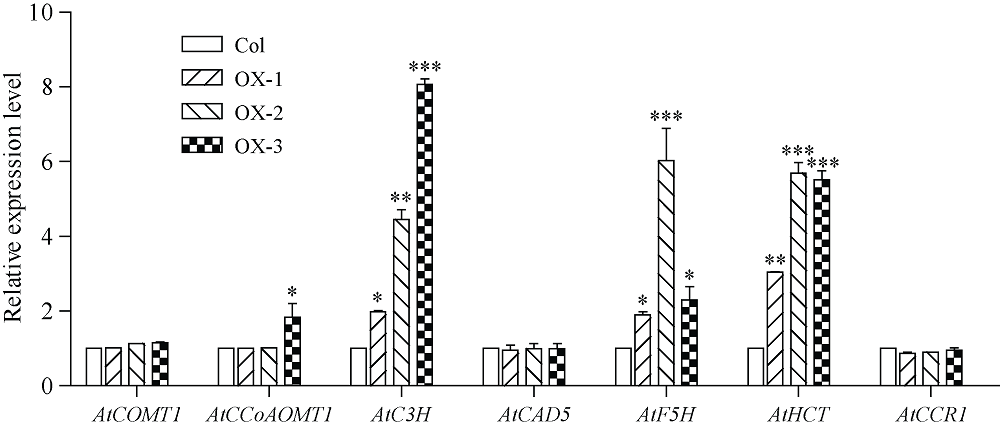
4-香豆酸辅酶A连接酶(4CL; EC 6.2.1.12)位于苯丙烷途径分支点的上游, 是苯丙烷代谢途径的核心酶, 可产生木质素、黄酮类等化合物, 这些化合物对植物的生长发育及环境适应性均具有重要作用。在双子叶植物中, 有关4CL的研究较多, 而在单子叶植物尤其是作物中的研究相对较少。本研究利用RACE技术从普通小麦中克隆了一个4CL基因Ta4CL1。系统发育分析表明, Ta4CL1与水稻、玉米和高粱等植物中在木质素合成中具有重要作用的4CLs聚成一类; 利用Ta4CL1过表达、拟南芥4CLs突变体at4cl1、at4cl3和at4cl14cl3及其功能回复株系进行的分析表明, Ta4CL1与At4CL1功能相似, 在植物木质素合成中具有重要作用, 但未参与黄酮类化合物生物合成的调控过程; Ta4CL1是转基因拟南芥中4CL酶活性的主要贡献者。过表达Ta4CL1的转基因拟南芥叶片增大、茎更粗; Ta4CL1的表达还受茉莉酸甲酯(Methyl jasmonic acid, MeJA)、赤霉素(Gibberellin, GA)和生长素(Indoleacetic acid, IAA)等激素处理的影响。本研究为利用基因工程将Ta4CL1应用于改善小麦秸秆的利用效率提供了理论依据。
| [1] | Chen F, Dixon R A. Lignin modification improves fermentable sugar yields for biofuel production. Nat Biotechnol, 2007, 25:759-761. |
| [2] | Tarasov D, Leitch M, Fatehi P. Lignin-carbohydrate complexes: properties, applications, analyses, and methods of extraction: a review. Biotechnol Biofuels, 2018, 11:269. |
| [3] | Boerjan W, Ralph J, Baucher M. Lignin biosynthesis. Annu Rev Plant Biol, 2003, 54:519-546. |
| [4] | Sun H, Guo K, Feng S, Zou W, Li Y, Fan C, Peng L. Positive selection drives adaptive diversification of the 4-coumarate: CoA ligase (4CL) gene in angiosperms. Ecol Evol, 2015, 5:3413-3420. |
| [5] | Lavhale S G, Kalunke R M, Giri A P. Structural, functional and evolutionary diversity of 4-coumarate-CoA ligase in plants. Planta, 2018, 248:1063-1078. |
| [6] | Li Y, Kim J I, Pysh L, Chapple C. Four isoforms ofArabidopsis 4-coumarate: CoA ligase have overlapping yet distinct roles in phenylpropanoid metabolism. Plant Physiol, 2015, 169:2409-2421. |
| [7] | Sun H, Li Y, Feng S, Zou W, Guo K, Fan C, Si S, Peng L. Analysis of five rice 4-coumarate: coenzyme A ligase enzyme activity and stress response for potential roles in lignin and flavonoid biosynthesis in rice. Biochem Biophys Res Commun, 2013, 430:1151-1156. |
| [8] | Xiong W, Wu Z, Liu Y, Li Y, Su K, Bai Z, Guo S, Hu Z, Zhang Z, Bao Y, Sun J, Yang G, Fu C. Mutation of 4-coumarate: coenzyme A ligase 1 gene affects lignin biosynthesis and increases the cell wall digestibility in maize brown midrib5 mutants. Biotechnol Biofuels, 2019, 12:82. |
| [9] | Rao G, Pan X, Xu F, Zhang Y, Cao S, Jiang X, Lu H. Divergent and overlapping function of five 4-coumarate/coenzyme A ligases from Populus tomentosa. Plant Mol Biol Rep, 2015, 33:841-885. |
| [10] | Hu W J, Kawaoka A, Tsai C J, Lung J, Osakabe K, Ebinuma H, Chiang V L. Compartmentalized expression of two structurally and functionally distinct 4-coumarate: CoA ligase genes in aspen (Populus tremuloides). Proc Natl Acad Sci USA, 1998, 95:5407-5412. |
| [11] | Hatfield R, Ralph J, Grabber J H. A potential role for sinapyl p-coumarate as a radical transfer mechanism in grass lignin formation. Planta, 2008, 228:919-928. |
| [12] | Gui J, Shen J, Li L. Functional characterization of evolutionarily divergent 4-coumarate: coenzyme a ligases in rice. Plant Physiol, 2011, 157:574-586. |
| [13] | Sattler S E, Funnell-Harris D L, Pedersen J F. Brown midrib mutations and their importance to the utilization of maize, sorghum, and pearl millet lignocellulosic tissues. Plant Sci, 2010, 178:229-238. |
| [14] | Saballos A, Sattler S E, Sanchez E, Foster T P, Xin Z, Kang C, Pedersen J F, Vermerris W. Brown midrib2 (Bmr2) encodes the major 4-coumarate: coenzyme A ligase involved in lignin biosynthesis in sorghum(Sorghum bicolor (L.) Moench). Plant J, 2012, 70:818-830. |
| [15] | Clough S J, Bent A F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J, 1998, 16:735-743. |
| [16] | 张双双, 王立伟, 姚楠, 郭光艳, 夏玉凤, 秘彩莉. 水稻OsUBA基因的表达及其在促进种子萌发和开花中的功能. 作物学报, 2019, 45:1327-1337. |
| Zhang S S, Wang L W, Yao N, Guo G Y, Xia Y F, Bei C L. Expression of OsUBA and its function in promoting seed germination and flowering. Acta Agron Sin, 2019, 45:1327-1337 (in Chinese with English abstract). | |
| [17] | Eudes A, Pollet B, Sibout R, Do C T, Séguin A, Lapierre C, Jouanin L. Evidence for a role of AtCAD1 in lignification of elongating stems of Arabidopsis thaliana. Planta, 2006, 225:23-39. |
| [18] | 薛应钰, 师桂英, 徐秉良, 陈荣贤. 美洲南瓜(Cucurbita pepo)种皮发育形态观察及其相关酶活性测定. 草业学报, 2011, 20(2):23-30. |
| Xue Y Y, Shi G Y, Xu B L, Chen R X. Studies on morphology of seed coat development and its related enzyme activity assay inCucurbita pepo. Acta Pratac Sin, 2011, 20(2):23-30 (in Chinese with English abstract). | |
| [19] | Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25:402-408. |
| [20] | Franke R, Humphreys J M, Hemm M R, Denault J W, Ruegger M O, Cusumano J C, Chapple C. The Arabidopsis REF8 gene encodes the 3-hydroxylase of phenylpropanoid metabolism. Plant J, 2002, 30:33-45. |
| [21] | Franke R, Hemm M R, Denault J W, Ruegger M O, Humphreys J M, Chapple C. Changes in secondary metabolism and deposition of an unusual lignin in the ref8 mutant of Arabidopsis. Plant J, 2002, 30:47-59. |
| [22] | Meyer K, Shirley A M, Cusumano J C, Bell-Lelong D A, Chapple C. Lignin monomer composition is determined by the expression of a cytochrome P450-dependent monooxygenase in Arabidopsis. Proc Natl Acad Sci USA, 1998, 95:6619-6623. |
| [23] | Besseau S, Hoffmann L, Geoffroy P, Lapierre C, Pollet B, Legrand M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell, 2007, 19:148-162. |
| [24] | Wagner A, Donaldson L, Kim H, Phillips L, Flint H, Steward D, Torr K, Koch G, Schmitt U, Ralph J. Suppression of 4-coumarate-CoA ligase in the coniferous gymnosperm Pinus radiata. Plant Physiol, 2009, 149:370-383. |
| [25] | Lee D, Douglas C J. Two divergent members of a tobacco 4-coumarate: coenzyme A ligase (4CL) gene family. cDNA structure, gene inheritance and expression, and properties of recombinant proteins. Plant Physiol, 1996, 112:193-205. |
| [26] | Gao S, Yu H N, Xu R X, Cheng A X, Lou H X. Cloning and functional characterization of a 4-coumarate CoA ligase from liverwort Plagiochasma appendiculatum. Phytochemistry, 2015, 111:48-58. |
| [27] | Bi C, Chen F, Jackson L, Gill B Sand Li W. Expression of lignin biosynthetic genes in wheat during development and upon infection by fungal pathogens. Plant Mol Biol Rep, 2011, 29:149-161. |
| [1] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [2] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [3] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [4] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
| [5] | 刘运景, 郑飞娜, 张秀, 初金鹏, 于海涛, 代兴龙, 贺明荣. 宽幅播种对强筋小麦籽粒产量、品质和氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 716-725. |
| [6] | 马红勃, 刘东涛, 冯国华, 王静, 朱雪成, 张会云, 刘静, 刘立伟, 易媛. 黄淮麦区Fhb1基因的育种应用[J]. 作物学报, 2022, 48(3): 747-758. |
| [7] | 徐龙龙, 殷文, 胡发龙, 范虹, 樊志龙, 赵财, 于爱忠, 柴强. 水氮减量对地膜玉米免耕轮作小麦主要光合生理参数的影响[J]. 作物学报, 2022, 48(2): 437-447. |
| [8] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [9] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
| [10] | 马博闻, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑, 姜东. 花前渍水锻炼调控花后小麦耐渍性的生理机制研究[J]. 作物学报, 2022, 48(1): 151-164. |
| [11] | 韦一昊, 于美琴, 张晓娇, 王露露, 张志勇, 马新明, 李会强, 王小纯. 小麦谷氨酰胺合成酶基因可变剪接分析[J]. 作物学报, 2022, 48(1): 40-47. |
| [12] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [13] | 罗江陶, 郑建敏, 蒲宗君, 范超兰, 刘登才, 郝明. 四倍体小麦与六倍体小麦杂种的染色体遗传特性[J]. 作物学报, 2021, 47(8): 1427-1436. |
| [14] | 王艳朋, 凌磊, 张文睿, 王丹, 郭长虹. 小麦B-box基因家族全基因组鉴定与表达分析[J]. 作物学报, 2021, 47(8): 1437-1449. |
| [15] | 习玲, 王昱琦, 朱微, 王益, 陈国跃, 蒲宗君, 周永红, 康厚扬. 78份四川小麦育成品种(系)条锈病抗性鉴定与抗条锈病基因分子检测[J]. 作物学报, 2021, 47(7): 1309-1323. |
|
||