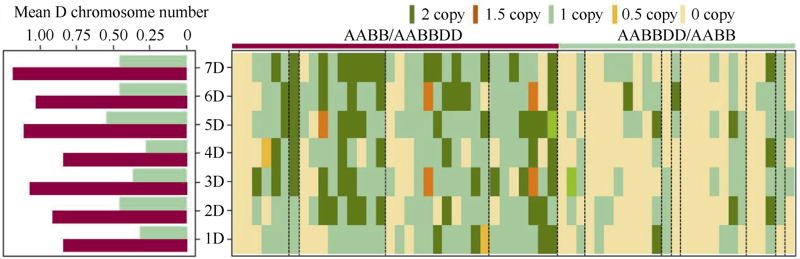
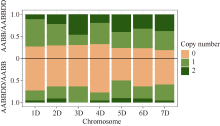
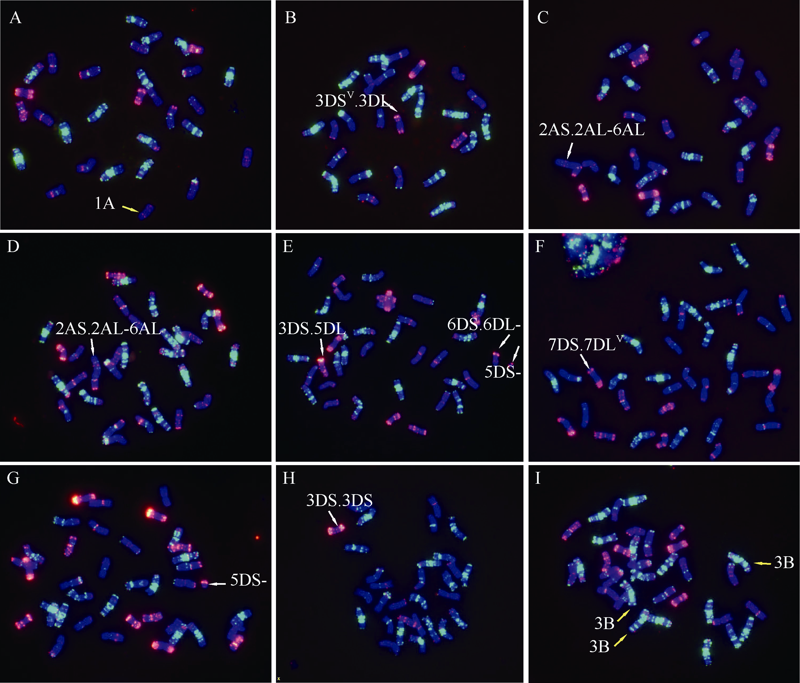
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2021, Vol. 47 ›› Issue (8): 1427-1436.doi: 10.3724/SP.J.1006.2021.01067
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
LUO Jiang-Tao1( ), ZHENG Jian-Min1, PU Zong-Jun1,*(), FAN Chao-Lan2, LIU Deng-Cai2, HAO Ming2,*()
), ZHENG Jian-Min1, PU Zong-Jun1,*(), FAN Chao-Lan2, LIU Deng-Cai2, HAO Ming2,*()
| [1] |
Shewry P R, Hey S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J Cereal Sci, 2015,65:236-243.
doi: 10.1016/j.jcs.2015.07.014 |
| [2] | Kihara H. Discovery of the DD-analyser, one of the ancestors of Triticum vulgare. Agric Hort, 1944,19:889-890. |
| [3] | McFadden E S, Sears E R. The artifcial synthesis of Triticum spelta. Rec Genet Soc Am, 1944,13:26-27. |
| [4] |
Huang S X, Sirikhachornkit A, Su X J, Faris J D, Gill B, Haselkorn R, Gornicki P. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc Natl Acad Sci USA, 2002,99:8133-8138.
doi: 10.1073/pnas.072223799 |
| [5] |
Klymiuk V, Yaniv E, Huang L, Raats D, Fatiukha A, Chen S S, Feng L H, Frenkel Z, Krugman T, Lidzbarsky G, Chang W, Jääskeläinen M J, Schudoma C, Paulin L, Laine P, Bariana H, Sela H, Saleem K, Sørensen C K, Hovmøller M S, Distelfeld A, Chalhoub B, Dubcovsky J, Korol A B, Schulman A H, Fahima T. Cloning of the wheat Yr15 resistance gene sheds light on the plant tandem kinase-pseudokinase family. Nat Commun, 2018,9:3735.
doi: 10.1038/s41467-018-06138-9 |
| [6] |
He Y, Feng L H, Jiang Y, Zhang L Q, Yan J, Zhao G, Wang J R, Chen G Y, Wu B H, Liu D C, Huang L, Fahima T. Distribution and nucleotide diversity of Yr15 in wild emmer populations and Chinese wheat germplasm. Pathogens, 2020,9:212.
doi: 10.3390/pathogens9030212 |
| [7] |
Li A L, Liu D C, Yang W Y, Kishii M, Mao L. Synthetic hexaploid wheat: yesterday, today, and tomorrow. Engineering, 2018,4:552-558.
doi: 10.1016/j.eng.2018.07.001 |
| [8] |
Hao M, Zhang L Q, Zhao L B, Dai S F, Li A L, Yang W Y, Xie D, Li Q C, Ning S Z, Yan Z H, Wu B H, Lan X J, Yuan Z W, Huang L, Wang J R, Zheng K, Chen W S, Yu M, Chen X J, Chen M P, Wei Y M, Zhang H G, Kishii M, Hawkesford M, Mao L, Zheng Y L, Liu D C. A breeding strategy targeting the secondary gene pool of bread wheat: introgression from a synthetic hexaploid wheat. Theor Appl Genet, 2019,132:2285-2294.
doi: 10.1007/s00122-019-03354-9 |
| [9] |
Dvorak J, Akhunov E D, Akhunov A R, Deal K R, Luo M C. Molecular characterization of a diagnostic DNA marker for domesticated tetraploid wheat provides evidence for gene flow from wild tetraploid wheat to hexaploid wheat. Mol Biol Evol, 2006,23:1386-1396.
pmid: 16675504 |
| [10] |
Cheng H, Liu J, Wen J, Nie X J, Xu L H, Chen N B, Li Z X, Wang Q L, Zheng Z Q, Li M, Cui L C, Bian J X, Wang Z H, Xu S B, Yang Q, Appels R, Han D J, Song W N, Sun Q X, Jiang Y. Frequent intra- and inter-species introgression shapes the landscape of genetic variation in bread wheat. Genome Biol, 2019,20:136.
doi: 10.1186/s13059-019-1744-x pmid: 31300020 |
| [11] |
He F, Pasam R, Shi F, Kant S, Keeble-Gagnere G, Kay P, Forrest K, Fritz A, Hucl P, Wiebe K, Knox R, Cuthbert R, Pozniak C, Akhunova A, Morrell P L, Davies J P, Webb S R, Spangenberg G, Hayes B, Daetwyler H, Tibbits J, Hayden M, Akhunov E. Exome sequencing highlights the role of wild-relative introgression in shaping the adaptive landscape of the wheat genome. Nat Genet, 2019,51:896-904.
doi: 10.1038/s41588-019-0382-2 |
| [12] |
Briggle L W. Transfer of resistance to Erysiphe graminis f. sp. tritici from Khapli emmer and Yuma durum to hexaploid wheat. Crop Sci, 1966,6:459-461.
doi: 10.2135/cropsci1966.0011183X000600050020x |
| [13] |
Reader S M, Miller T E. The introduction into bread wheat of a major gene for resistance to powdery mildew from wild emmer wheat. Euphytica, 1991,53:57-60.
doi: 10.1007/BF00032033 |
| [14] |
Rong J K, Millet E, Manisterski J, Feldman M. A new powdery mildew resistancegene: Introgression from wild emmer into common wheat and RFLP-based mapping. Euphytica, 2000,115:121-126.
doi: 10.1023/A:1003950431049 |
| [15] |
Liu Z Y, Sun Q X, Ni Z F, Nevo E, Yang T. Molecular characterization of a novel powdery mildew resistance gene Pm30 in wheat originating from wild emmer. Euphytica, 2002,123:21-29.
doi: 10.1023/A:1014471113511 |
| [16] | 张连松, 华为, 关海英, 李根桥, 张宏涛, 解超杰, 杨作民, 孙其信, 刘志勇. 野生二粒小麦导入普通小麦的抗白粉病基因MlWE29分子标记定位. 作物学报, 2009,35:998-1005. |
| Zhang L S, Hua W, Guan H Y, Li G Q, Xie C J, Yang Z M, Sun Q X, Liu Z Y. Molecular mapping of powdery mildew resistance gene MIWE29 in wheat originated from wild emmer (Triticum turgidum var. dicoccoides). Acta Agron Sin, 2009,35:998-1005 (in Chinese with English abstract). | |
| [17] | 解超杰, 倪中福, 孙其信, 杨作民, 刘保申, 魏艳玲. 利用小麦微卫星标记定位一个来自野生二粒小麦的抗白粉病基因. 遗传学报, 2001,28:1034-1039. |
| Xie C J, Ni Z F, Sun Q X, Yang Z M, Liu B K, Wei Y L. Molecular tagging of a major powdery mildew resistance gene MlG in wheat derived from wild emmer by using microsatellite maker. Acta Genet Sin, 2001,28:1034-1039 (in Chinese with English abstract) | |
| [18] |
Mesfin A, Frohberg R C, Anderson J A. RFLP markers associated with high grain protein from Triticum turgidum L. dicoccoides introgressed into hard red spring wheat. Crop Sci, 1999,39:508-513.
doi: 10.2135/cropsci1999.0011183X003900020035x |
| [19] |
Lanning S P, Blake N K, Sherman J D, Talbert L E. Variable production of tetraploid and hexaploid progeny lines from spring wheat by durum wheat crosses. Crop Sci, 2008,48:199-202.
doi: 10.2135/cropsci2007.06.0334 |
| [20] |
Eberhard F S, Zhang P, Lehmensiek A, Hare R A, Simpfendorfer S, Sutherland M W. Chromosome composition of an F2 Triticum aestivum × T. turgidum spp. durum cross analyzed by DArT markers and MCFISH. Crop Pasture Sci, 2010,61:619-624.
doi: 10.1071/CP10131 |
| [21] |
Martin A, Simpfendorfer S, Hare R A, Eberhard F S, Sutherland M W. Retention of D genome chromosomes in pentaploid wheat crosses. Heredity, 2011,107:315-319.
doi: 10.1038/hdy.2011.17 pmid: 21427754 |
| [22] |
Kalous J R, Martin J M, Sherman J D, Heo H Y, Blake N K, Lanning S P, Eckhoff J L A, Chao S, Akhunov E, Talbert L E. Impact of the D genome and quantitative trait loci on quantitative traits in a spring durum by spring bread wheat cross. Theor Appl Genet, 2015,128:1799-1811.
doi: 10.1007/s00122-015-2548-3 |
| [23] |
Ren R S, Ray R, Li P F, Xu J H, Zhang M, Liu G, Yao X F, Kilian A, Yang X P. Construction of a high-density DArTseq SNP-based genetic map and identification of genomic regions with segregation distortion in agenetic population derived from a cross between feral and cultivated-type watermelon. Mol Genet Genomics, 2015,290:1457-1470.
doi: 10.1007/s00438-015-0997-7 |
| [24] |
Padmanaban S, Sutherland M W, Knight N L, Martin A. Genome inheritance in populations derived from hexaploid/tetraploid and tetraploid/hexaploid wheat crosses. Mol Breed, 2017,37:48.
doi: 10.1007/s11032-017-0647-3 |
| [25] |
Padmanaban S, Zhang P, Hare R A, Sutherland M W, Martin A. Pentaploid wheat hybrids: applications, characterisation, and challenges. Front Plant Sci, 2017,8:358.
doi: 10.3389/fpls.2017.00358 pmid: 28367153 |
| [26] |
Padmanaban S, Zhang P, Sutherland M W, Knight N L, Martin A. A cytological and molecular analysis of D-genome chromosome retention following F2-F6 generations of hexaploid × tetraploid wheat crosses. Crop Pasture Sci, 2018,69:121-130.
doi: 10.1071/CP17240 |
| [27] |
Schwarzacher T, Leitch A R, Bennett M D, Heslop-Harrison J S. In situ localization of parental genomes in a wide hybrid. Ann Bot-London, 1989,64:315-324.
doi: 10.1093/oxfordjournals.aob.a087847 |
| [28] |
Lim K B, Ramanna M, Jacobsen E, van Tuyl J. Evaluation of BC2 progenies derived from 3 x-2x and 3x-4x crosses of Lilium hybrids: a GISH analysis. Theor Appl Genet, 2003,106:568-574.
doi: 10.1007/s00122-002-1070-6 |
| [29] |
Huang X Y, Zhu M Q, Zhuang L F, Zhang S Y, Wang J J, Chen X Y, Wang D R, Chen J Y, Bao Y G, Guo J, Zhang J L, Feng Y G, Chu C G, Du P, Qi Z J, Wang H G, Chen P D. Structural chromosome rearrangements and polymorphisms identified in Chinese wheat cultivars by high-resolution multiplex oligonucleotide FISH. Theor Appl Genet, 2018,131:1967-1986.
doi: 10.1007/s00122-018-3126-2 |
| [30] |
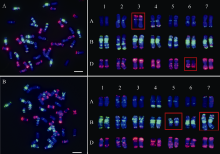
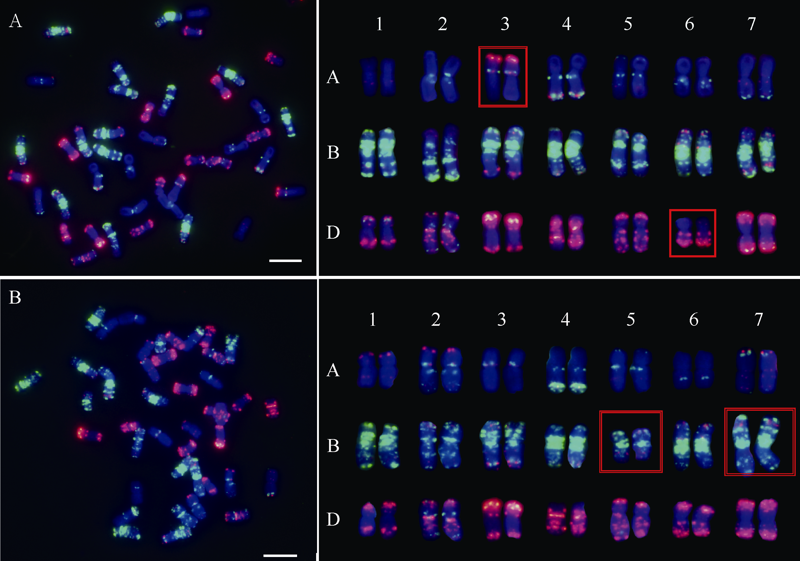
Luo J T, Zhao L B, Zheng J M, Li Y Z, Zhang L Q, Liu D C, Pu Z J, Hao M. Karyotype mosaicism in early generation synthetic hexaploid wheats. Genome, 2020,63:329-336.
doi: 10.1139/gen-2019-0148 |
| [31] |
Du P, Zhuang L F, Wang Y Z, Yuan L, Wang Q, Dawadondup, Wang D R, Tan L J, Shen J, Xu H B, Zhao H, Chu C G, Qi Z J. Development of oligonucleotides and multiplex probes for quick and accurate identification of wheat and Thinopyrum bessarabicum chromosomes. Genome, 2017,60:93-103.
doi: 10.1139/gen-2016-0095 |
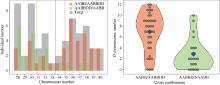
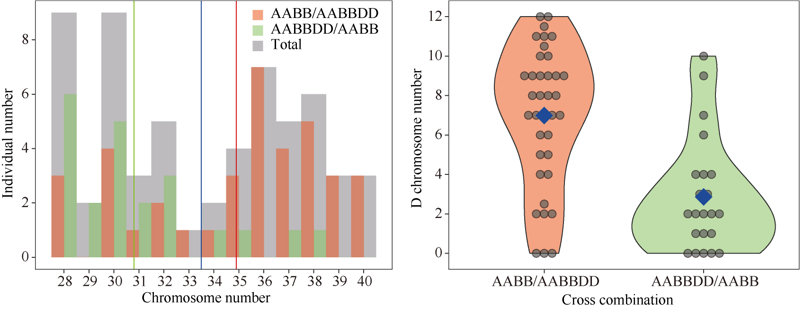
| [32] | Wickham H. Ggplot2: Elegant Graphics for Data Analysis. New York, NY: Springer-Verlag, 2016. |
| [33] |
Delhaize E, James R A, Ryan P R. Aluminium tolerance of root hairs underlies genotypic differences in Rhizosheath size of wheat (Triticum aestivum) grown on acid soil. New Phytol, 2012,195:609-619.
doi: 10.1111/nph.2012.195.issue-3 |
| [34] |
Dvořák J, Gorham J. Methodology of gene transfer by homoeologous recombination into Triticum turgidum: transfer of K+/Na+ discrimination from Triticum aestivum. Genome, 1992,35:639-646.
doi: 10.1139/g92-096 |
| [35] |
Luo M C, Dubcovsky J, Goyal S, Dvořáket J. Engineering of interstitial foreign chromosome segments containing the K+/Na+ selectivity gene Kna1 by sequential homoeologous recombination in durum wheat. Theor Appl Genet, 1996,93:1180-1184.
doi: 10.1007/BF00230144 pmid: 24162500 |
| [36] |
Han C, Zhang P, Ryan P R, Rathjen T M, Yan Z H, Delhaize E. Introgression of genes from bread wheat enhances the Aluminium tolerance of durum wheat. Theor Appl Genet, 2016,129:729-739.
doi: 10.1007/s00122-015-2661-3 |
| [37] |
Liu J, Huang L, Wang C Q, Liu Y X, Yan Z H, Wang Z Z, Xiang L, Zhong X Y, Gong F Y, Zheng Y L, Liu D C, Wu B H. Genome-wide association study reveals novel genomic regions associated with high grain protein content in wheat lines derived from wild emmer wheat. Front Plant Sci, 2019,10:464.
doi: 10.3389/fpls.2019.00464 |
| [38] |
Xiang L, Huang L, Gong F Y, Liu J, Wang Y F, Jin Y R, He Y, He J S, Jiang Q T, Zheng Y L, Liu D C, Wu B H. Enriching LMW-GS alleles and strengthening gluten properties of common wheat through wide hybridization with wild emmer. 3 Biotech, 2019,9:355.
doi: 10.1007/s13205-019-1887-1 pmid: 31501756 |
| [39] | Kihara H. Wheat Studies: Retrospect and Prospects (Developments in Crop Science). Tokyo: Kodansha Ltd, 1982. |
| [40] |
Sharma H C, Gill B S. Current status of wide hybridization in wheat. Euphytica, 1983,32:17-31.
doi: 10.1007/BF00036860 |
| [41] |
Friebe B, Zhang P, Linc G, Gill B S. Robertsonian translocations in wheat arise by centric misdivision of univalents at anaphase I and rejoining of broken centromeres during interkinesis of meiosis II. Cytogenet Genome Res, 2005,109:293-297.
doi: 10.1159/000082412 |
| [1] | LI Chao-Su,WU Xiao-Li,TANG Yong-Lu,LI Jun,MA Xiao-Ling,LI Shi-Zhao,HUANG Ming-Bo,LIU Miao. Response of yield and associated physiological characteristics for different wheat cultivars to nitrogen stress at mid-late growth stage [J]. Acta Agronomica Sinica, 2019, 45(8): 1260-1269. |
| [2] | LI Jun, WEI Hui-Ting, HU Xiao-Rong, LI Chao-Su, TANG Yong-Lu, LIU De-Cai, YANG Wu-Yun. Identification of a High-Yield Introgression Locus from Synthetic Hexaploid Wheat in Chuanmai 42 [J]. Acta Agron Sin, 2011, 37(02): 255-262. |
| [3] | LI Jun,WEI Hui-Ting,YANG Su-Jie,LI Chao-Su,TANG Yong-Lu,HU Xiao-Rong,YANG Wu-Yun. Genetic Effects of 1BS Chromosome Arm on the Main Agrionomic Traits in Chuanmai 42 [J]. Acta Agron Sin, 2009, 35(12): 2167-2173. |
| [4] | LIAO Jie;WEI Hui-Ting;LI Jun;YANG Yu-Min;ZENG Yun-Chao;PENG Zheng-Song;YANG Wu-Yun. Detection of the Introgression Loci of Synthetic Hexaploid Wheat in Wheat Cultivar Chuanmai 42 by SSR Markers [J]. Acta Agron Sin, 2007, 33(05): 703-707. |
| [5] | LI Gen-Ying;XIA Xian-Chun;ZHANG Ming;ZHANG Yong;HE Zhong-Hu;SUN Qi-Xin. Allelic Variations of Puroindoline a and Puroindoline b Genes in New Type of Synthetic Hexaploid Wheats from CIMMYT [J]. Acta Agron Sin, 2007, 33(02): 242-249. |
|
||