欢迎访问作物学报,今天是
作物学报 ›› 2018, Vol. 44 ›› Issue (11): 1733-1742.doi: 10.3724/SP.J.1006.2018.01733
• 研究简报 • 上一篇
邢芦蔓,吕伟增,雷薇,梁雨欢,卢洋,陈军营( )
)
Lu-Man XING,Wei-Zeng LYU,Wei LEI,Yu-Huan LIANG,Yang LU,Jun-Ying CHEN()
摘要:
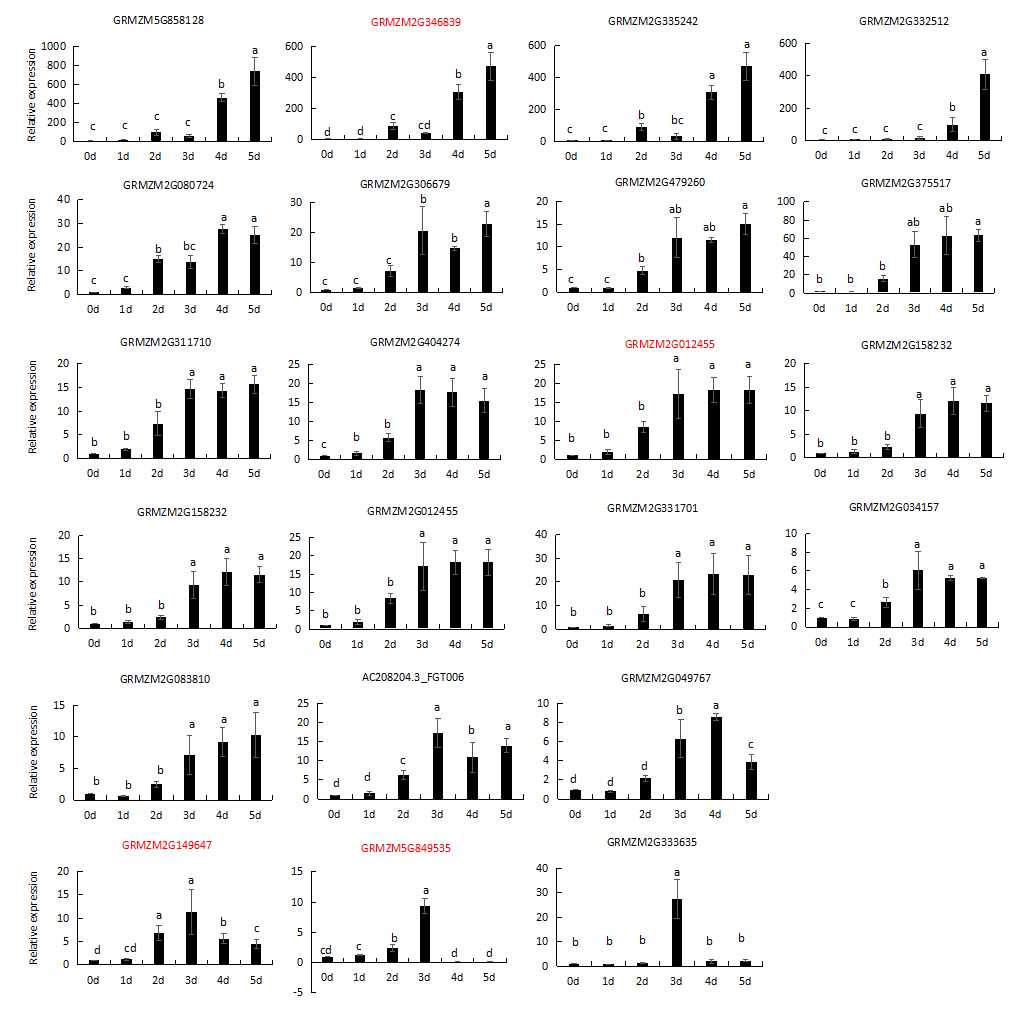
以玉米杂交种“郑单958”为材料, 采用高温(45°C)高湿(100%相对湿度)对玉米种子进行人工老化处理, 并用转录组技术, 研究植物HSP20基因对种子人工老化处理的响应, 旨在为揭示种子衰老的分子机制提供依据。结果表明, 随着老化时间的延长, 种子活力和种胚内过氧化氢酶的活性均表现下降趋势; 过氧化氢含量在老化第3天达到最大值, 随后下降; 丙二醛含量逐渐升高; 转录组检测表明, 种子老化过程中差异显著的HSP20基因有25个, 这些基因编码的HSP20蛋白主要被定位在细胞核、线粒体、以及叶绿体上, 其序列中均含有ACD保守序列(RVDWRETPDAHEIVVDVP GMRREDLRIEVEDNRVLRVSGERRRAEERKGDH WHREERSYGRFWRRFRLPENADLDSVAASLDSGVL TVRFRK)。该序列中含有较多的Arg (11.2%)、Lys (7.2%)、Pro (4.2%)、Thr (3.9%)等氨基酸, 老化过程中积累的ROS可能氧化这些氨基酸, 导致HSP20结构破坏、功能丧失。利用qRT-PCR技术对挑选的编码细胞质、叶绿体和线粒体HSP20的基因的表达模式分析显示, 随着老化程度的加深, 2个编码细胞质HSP20的基因上调表达, 另外两个编码叶绿体和线粒体HSP20基因的表达量在老化第3天达最大值, 随后下降。推测HSP20基因对种子老化有重要作用, HSP20蛋白的ACD结构域中Arg、Lys等氨基酸的靶向氧化可能是种子衰老的主要原因之一。
| [1] |
Mohamed H , Al-Whaibi. Plant heat- shock protein: a mini review. J King Saud Univ Sci, 2011,23:139-150
doi: 10.1016/j.jksus.2010.06.022 |
| [2] |
Joshi C P, Nguyen H T . Differential display-mediated rapid identification of different members of a multigene family, HSP16.9 in wheat. Plant Mol Biol, 1996,31:575-584
doi: 10.1007/BF00042230 |
| [3] |
Kyeong K K, Rosalind K, Sung H K . Crystal structure of a small heat-shock protein. Nature, 1998,394:595-599
doi: 10.1038/29106 pmid: 9707123 |
| [4] |
Haslbeck M . sHsps and their role in the chaperone network. Cell Mol Life Sci, 2002,59:1649-1657
doi: 10.1007/PL00012492 pmid: 12475175 |
| [5] |
Franck E , Madsen O, van Rheede T, Ricard G, Huynen M A, de Jong W W. Evolutionary diversity of vertebrate small heat shock proteins. J Mol Evol, 2004,59:792-805
doi: 10.1007/s00239-004-0013-z |
| [6] | Seo J S, Lee Y M, Park H G, Lee J S . The intertidal copepod Tigriopus japonicus small heat shock protein 20 gene( Hsp20) enhances thermotolerance of transformed Escherichia coli. Biochem Biophys Res Commun, 2006,340:901-908 |
| [7] |
Waters E R, Lee G J, Vierling E . Evolution, structure and function of the small heat shock proteins in plants. J Exp Bot, 1996,47:325-338
doi: 10.1093/jxb/47.3.325 |
| [8] | Helm K W, Schmeits J, Vierling E . An endomembrane-localized small heat-shock protein fromArabidopsis thaliana.Plant Physiol, 1995,107:287-288 |
| [9] |
Vierling E . The roles of heat shock proteins in plants. Annu Rev Plant Biol, 1991,42:579-620
doi: 10.1146/annurev.pp.42.060191.003051 |
| [10] | Scharf K D, Siddique M, Vierling E . The expanding family ofArabidopsis thaliana small heat stress proteins and a new family of proteins containing alpha-crystallin domains (Acd proteins). Cell Stress Chaperones, 2001,6:225-237 |
| [11] |
LaFayette P R, Nagao R T, O’Grady K, Vierling E, Key J L . Molecular characterization of cDNAs encoding low-molecular- weight heat shock proteins of soybean. Plant Mol Biol, 1996,30:159-169
doi: 10.1007/BF00017810 |
| [12] |
Basha E, Friedrich K L, Vierling E . The N-terminal arm of small heat shock proteins is important for both chaperone activity and substrate specificity. J Biol Chem, 2006,281:39943-39952
doi: 10.1074/jbc.M607677200 |
| [13] |
Jiao W W, Li P L, Zhang J R, Zhang H, Chang Z Y . Small heat-shock proteins function in the insoluble protein complex. Biochem Biophys Res Commun, 2005,335:227-231
doi: 10.1016/j.bbrc.2005.07.065 pmid: 16055090 |
| [14] |
Prieto-Dapena P, Castano R, Almoguera C, Jordano J . Improved resistance to controlled deterioration in transgenic seeds. Plant Physiol, 2006,142:1102-1112
doi: 10.1104/pp.106.087817 |
| [15] | Guo M, Liu J H, Lu J P, Zhai Y F, Wang H, Gong Z H, Wang S B, Lu M H . Genome-wide analysis of theCaHsp20 gene family in pepper: comprehensive sequence and expression profile analysis under heat stress. Front Plant Sci, 2015,6:806 |
| [16] | Zhou Y L, Chen H H, Chu P, Li Y, Tan B, Ding Y , Tsang E W T, Jiang L, Wu K, Huang S Z. NnHSP17.5, a cytosolic class II small heat shock protein gene from Nelumbo nucifera, contributes to seed germination vigor and seedling the rmotolerance in transgenic Arabidopsis. Plant Cell Rep, 2012,2:379-389 |
| [17] | Kaur H, Petla B P, Kamble N U, Singh A, Rao V . Differentially expressed seed aging responsive heat shock protein OsHSP18.2 implicates in seed vigor, longevity and improves germination and seedling establishment under abiotic stress. Front Plant Sci, 2015,6:713 |
| [18] |
Chauhan H, Khurana N, Nijhavan A, Khurana J P, Khurana P . The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ, 2012,35:1912-1931
doi: 10.1111/j.1365-3040.2012.02525.x |
| [19] |
Muthusamy S K, Dalal M, Chinnusamy V , Bansal B K C. Genome-wide identification and analysis of biotic and abiotic stress regulation of small heat shock protein (HSP20) family genes in bread wheat. J Plant Physiol, 2017,211:100-113
doi: 10.1016/j.jplph.2017.01.004 |
| [20] | Sun L P, Liu Y, Kong X P, Zhang D, Pan J W, Zhou Y, Wang L, Li D Q, Yang X H . ZmHSP16.9, a cytosolic class I small heat shock protein in maize (Zea mays ), confers heat tolerance in transgenic tobacco. Plant Cell Rep, 2012,31:1473-1484 |
| [21] | 孙爱清, 葛淑娟, 董伟, 单晓笛, 董树亭, 张杰道 . 玉米小分子热激蛋白ZmHSP17.7 基因的克隆与功能分析. 作物学报, 2015,41:414-421 |
| Sun A Q, Ge S J, Dong W, Shan X D, Dong S T, Zhang J D . Cloning and function analysis of small heat shock protein gene ZmHSP17.7 from maize. Acta Agron Sin, 2015,41:414-421 (in Chinese with English abstract) | |
| [22] | Hu X L, Yang Y F, Gong F P, Zhang D Y, Zhang L, Wu L J, Li C H, Wang W . Protein sHSP26 improves chloroplast performance under heat stress by interacting with specific chloroplast proteins in maize (Zea mays ). J Proteomics, 2015,115:81-92 |
| [23] | Basak O, Demir I, Mavi K. Matthews S . Controlled deterioration test for predicting seedling emergence and longevity of pepper (Capsicum annuum L.) seed lots. Seed Sci Technol, 2006,34:701-712 |
| [24] | Zhang L, Qiu Z, Hu Y, Yang F, Yan S, Zhao L, Li B, He S, Huang M, Li J, Li L . ABA treatment of germinating maize seeds inducesVP1 gene expression and selective promoter-associated histone acetylation. Physiol Plant, 2011,143:287-296 |
| [25] |
Jana S, Choudhur M A . Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquat Bot, 1982,12:345-354
doi: 10.1016/0304-3770(82)90026-2 |
| [26] | 李合生 . 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000. pp 164- 169, 260-261 |
| Li H S. Experimental Principles and Techniques of Plant Physiological Biochemical. Beijing: Higher Education Press, 2000. pp 164- 169, 260-261(in Chinese) | |
| [27] | 曹广灿, 林一欣, 薛梅真, 邢芦蔓, 吕伟增, 杨伟飞, 陈军营 . 玉米种胚内质网胁迫相关基因对人工老化处理的响应. 中国农业科学, 2016,49:429-442 |
| Cao G C, Lin Y X, Xue M Z, Xing L M, Lyu W Z, Yang W F, Chen J Y . Responses of endoplasmic reticulum stress-related genes in maize embryo to artificial aging treatment. Sci Agric Sin, 2016,49:429-442 (in Chinese with English abstract) | |
| [28] |
Basha E, Neill H O, Vierling E . Small heat shock proteins and alpha-crystallins: dynamic proteins with flexible functions. Trends Biochem Sci, 2012,37:106-117
doi: 10.1016/j.tibs.2011.11.005 pmid: 3460807 |
| [29] |
Levine R L, Garland D, Oliver C N, Amici A, Climent I, Lenz A G, Ahn B W, Shaltiel S, Stadtman E R . Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol, 1990,186:464-478
doi: 10.1016/0076-6879(90)86141-H |
| [30] | Siddique M, Gernhard S , Koskull-Döring P V, Vierling E, Scharf K D. The plant sHSP superfamily: five new members inArabidopsis thaliana with unexpected properties. Cell Stress Chaperones, 2008,13:183-197 |
| [31] |
Aung U T , McDonald M B. Changes in esterase activity associated with peanut (Arachis hypogeal L.) seed deterioration. Seed Sci Technol, 1995,23:101-111
doi: 10.1007/BF01276928 |
| [32] | Begnami C N, Cortelazzo A L . Cellular alterations during accelerated aging of French bean seeds. Seed Sci Technol, 1996,24:295-303 |
| [33] | Thapliyal R C, Connor K F . Effects of accelerated aging on viability, leachate exudation, and fatty acid content ofDalbergia sisso Roxb. Seed Sci Technol, 1997,25:311-319 |
| [34] | Rajjou L, Lovigny Y, Groot S P, Belghazi M, Job C, Job D . Proteome-wide characterization of seed aging in Arabidopsis : a comparison between artificial and natural aging protocols.Plant Physiol, 2008,148:620-641 |
| [35] | Lee G J, Pokala N, Vierling E . Structure and in vitro molecular chaperone activity of cytosolic small heat shock proteins from pea. J Biol Chem, 1995,270:10432-10438 |
| [36] |
Kim K H, Alam I, Kim Y G, Sharmin S A, Lee K W, Lee S H, Lee B H . Overexpression of a chloroplast-localized small heat shock protein OsHSP26 confers enhanced tolerance against oxidative and heat stresses in tall fescue. Biotechnol Lett, 2012,34:371-377
doi: 10.1007/s10529-011-0769-3 |
| [37] |
Lee B H, Won S H, Lee H S, Miyao M, Chung W I, Kim I J, Jo J . Expression of the chloroplast-localized small heat shock protein by oxidative stress in rice. Gene, 2000,245:283-290
doi: 10.1016/S0378-1119(00)00043-3 pmid: 10717479 |
| [38] |
Hamilton E W, Heckathorn S A . Mitochondrial adaptations to NaCl. Complex I is protected by anti-oxidants and small heat shock proteins, whereas complex II is protected by proline and betaine. Plant Physiol, 2001,126:1266-1274
doi: 10.1104/pp.126.3.1266 |
| [39] | Lund A A, Blum P H, Bhattramakki D, Elthon T E . Heat-stress response of maize mitochondria. Plant Physiol, 1998, 116:1097-1110 |
| [40] |
Smykal P, Masin J, Hrdy I, Konopasek I, Zarsky V . Chaperone activity of tobacco HSP18, a small heat-shock protein, is inhibited by ATP. Plant J, 2000,23:703-713
doi: 10.1046/j.1365-313x.2000.00837.x pmid: 10998182 |
| [41] | Lee S H, Lee K W, Lee D G, Son D, Park S J, Kim K Y, Park H S, Cha J Y . Identification and functional characterization of Siberian wild rye (Elymus sibiricus L.) small heat shock protein 16.9 gene( EsHsp16.9) conferring diverse stress tolerance in prokaryotic cells. Biotechnol Lett, 2015,37:881-890 |
| [42] |
Kim D H, Xu Z Y, Hwang I . AtHSP17.8 overexpression in transgenic lettuce gives rise to dehydration and salt stress resistance phenotypes through modulation of ABA-mediated signaling. Plant Cell Rep, 2013,32:1953-1963
doi: 10.1007/s00299-013-1506-2 |
| [43] |
Coca M A, Thomas T L, Jordano J . Differential regulation of small heat-shock genes in plants: analysis of a water-stress- inducible and developmentally activated sunflower promoter. Plant Mol Biol, 1996,31:863-876
doi: 10.1007/BF00019473 |
| [44] |
Chauhan H, Khurana N, Nijhavan A, Khurana J P, Khurnan P . The wheat chloroplastic small heat shock protein (sHSP26) is involved in seed maturation and germination and imparts tolerance to heat stress. Plant Cell Environ Plant, 2012,35:1912-1931
doi: 10.1111/j.1365-3040.2012.02525.x |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [8] | 李阿立, 冯雅楠, 李萍, 张东升, 宗毓铮, 林文, 郝兴宇. 大豆叶片响应CO2浓度升高、干旱及其交互作用的转录组分析[J]. 作物学报, 2022, 48(5): 1103-1118. |
| [9] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [10] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [11] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [12] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [13] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [14] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [15] | 张倩, 韩本高, 张博, 盛开, 李岚涛, 王宜伦. 控失尿素减施及不同配比对夏玉米产量及氮肥效率的影响[J]. 作物学报, 2022, 48(1): 180-192. |
|
||