欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (5): 827-836.doi: 10.3724/SP.J.1006.2021.02044
文钦( ), 贾思思, 王加峰, 黄翠红, 王慧, 陈志强, 郭涛*()
), 贾思思, 王加峰, 黄翠红, 王慧, 陈志强, 郭涛*()
WEN Qin(), JIA Si-Si, WANG Jia-Feng, HUANG Cui-Hong, WANG Hui, CHEN Zhi-Qiang, GUO Tao*()
摘要:
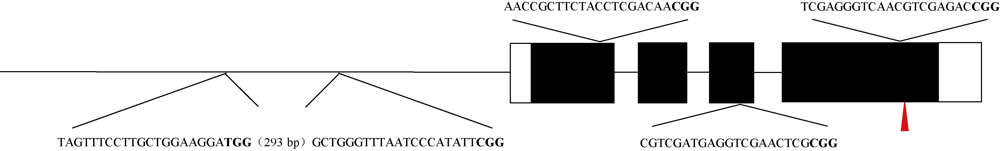
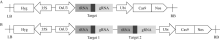
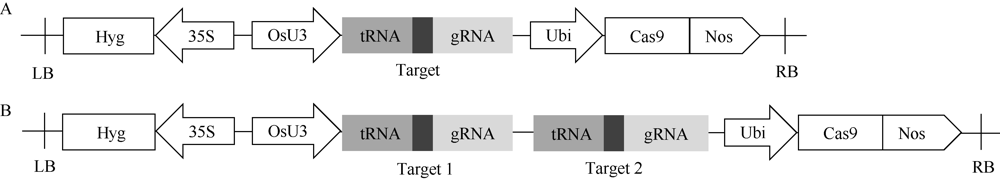
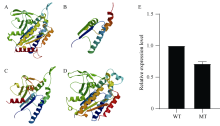
MTL基因是玉米中控制单倍体诱导性状的关键基因, 该基因在作物中的功能高度保守。OsMATL是MTL在水稻中的同源基因, 已有研究证实该基因的突变可以诱导水稻单倍体产生, 但不同遗传背景下, OsMATL基因的突变效应有待明确。本研究以日本晴(粳稻)和华航48 (籼稻)为材料, 利用CRISPR/Cas9技术对OsMATL基因启动子区和编码区的不同位点进行编辑, 成功获得OsMATL基因不同遗传背景系列突变体。分析了不同突变位点对结实率的影响, 发现OsMATL基因编码区突变后结实率为2%~15%, 且存在不同类型的败育籽粒, 而启动子区域的大片段缺失对结实率没有显著影响; 此外, 籼稻背景突变体的结实率高于粳稻突变体。本研究创制的不同类型OsMATL突变体为进一步研究水稻孤雌生殖单倍体诱导的机制提供了基础材料。
| [1] | 许阳东, 朱宽宇, 章星传, 王志琴, 杨建昌. 绿色超级稻品种的农艺与生理性状分析. 作物学报, 2019,45:70-80. |
| Xu Y D, Zhu K Y, Zhang X C, Wang Z Q, Yang J C. Analysis in agronomic and physiological traits of green super rice. Acta Agron Sin, 2019,45:70-80 (in Chinese with English abstract) | |
| [2] | Dunwell J M. Haploids in flowering plants: origins and exploitation. Plant Biotechnol J, 2010,8:377-424. |
| [3] | Chase S S. Production of homozygous diploids of maize from monoploids. Agron J, 1952,44:263-267. |
| [4] | 胡建林, 周黎, 郑兴飞, 董华林, 费震江, 查中萍, 游艾青, 徐得泽. 水稻花药离体培养的研究现状与展望. 农业科技通讯, 2019, (12):57-61. |
| Hu J L, Zhou L, Zheng, X F, Dong H L, Fei Z J, Cha Z P, You A Q, Xu D S. Research advance in twin-seedling in rice. Bull Agric Sci Tech, 2019, (12):57-61 (in Chinese with English abstract). | |
| [5] | 陈绍江. 作物育种工程化与工程化育种思考. 作物杂志, 2013, (6):1-4. |
| Chen S J. Crop breeding engineering and engineered breeding. Crops, 2013, (6):1-4 (in Chinese with English abstract). | |
| [6] | Prigge V, Xu X W, Li L, Babu R, Chen S J, Atlin G N, Melchinger A E. New insights into the genetics of in vivo induction of maternal haploids, the backbone of doubled haploid technology in maize. Genetics, 2012,190:781-793. |
| [7] | Kelliher T, Starr D, Richbourg L, Chintamanani S, Delzer B, Nuccio M L, Green J, Chen Z Y, McCuiston J, Wang W L, Liebler T, Bullock P, Martin B. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature, 2017,542:105-109. |
| [8] | Liu C X, Li X, Meng D X, Zhong Y, Chen C, Dong X, Xu X W, Chen B J, Li W, Li L, Tian X L, Zhao H M, Song W B, Luo H S, Zhang Q H, Lai J S, Jin W W, Yan J B, Chen S J. A 4-bp insertion at ZmPLA1 encoding a putative phospholipase a generates haploid induction in maize. Mol Plant, 2017,10:520-522. |
| [9] | Gilles L M, Khaled A, Laffaire J B, Chaignon S, Gendrot G, Laplaige J, Berges H, Beydon G, Bayle V, Barret P, Comadran J, Martinant J P, Rogowsky P M, Widiez T. Loss of pollen-specific phospholipase NOT LIKE DAD triggers gynogenesis in maize. EMBO J, 2017,36:707-717. |
| [10] | Dong L, Li L, Liu C L, Liu C X, Geng S F, Li X H, Huang C L, Mao L, Chen S J, Xie C X. Genome editing and double- fluorescence proteins enable robust maternal haploid induction and identification in maize. Mol Plant, 2018,11:1214-1217. |
| [11] | Yao L, Zhang Y, Liu C X, Liu Y B, Wang Y L, Liang D W, Liu J T, Sahoo G, Kelliher T. OsMATL mutation induces haploid seed formation in indica rice. Nat Plants, 2018,4:530-533. |
| [12] | Wang C, Liu Q, Shen Y, Hua Y F, Wang J J, Lin J R, Wu M G, Sun T T, Cheng Z K, Mercier R, Wang K J. Clonal seeds from hybrid rice by simultaneous genome engineering of meiosis and fertilization genes. Nat Biotechnol, 2019,37:283-286. |
| [13] | Liu C X, Zhong Y, Qi X L, Chen M, Liu Z K, Chen C, Tian X L, Li J L, Jiao Y Y, Wang D, Wang Y W, Li M R, Xin M M, Liu W X, Jin W W, Chen S J. Extension of the in vivo haploid induction system from diploid maize to hexaploid wheat. Plant Biotechnol J, 2020,18:316-318. |
| [14] | Liu H Y, Wang K, Jia Z M, Gong Q, Lin Z S, Du L P, Pei X W, Ye X G. Efficient induction of haploid plants in wheat by editing of TaMTL using an optimized Agrobacterium-mediated CRISPR system. J Exp Bot, 2019,71:1337-1349. |
| [15] | Madeira F, Park Y M, Lee J, Buso N, Gur T, Madhusoodanan N, Basutkar P, Tivey A R N, Potter S C, Finn R D, Lopez R. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res, 2019,47:W636-W641. |
| [16] | Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouzé P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res, 2002,30:325-327. |
| [17] | Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, Heer F T, de Beer T A P, Rempfer C, Bordoli L, Lepore R, Schwede T. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res, 2018,46:W296-W303. |
| [18] | Xie X R, Ma X L, Zhu Q L, Zeng D C, Li G S, Liu Y G. CRISPR-GE: a convenient software toolkit for CRISPR-based genome editing. Mol Plant, 2017,10:1246-1249. |
| [19] | Xie K B, Minkenberg B, Yang Y N. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA processing system. Proc Natl Acad Sci USA, 2015,112:3570-3575. |
| [20] | Hiei Y, Ohta S, Komari T, Kumashiro T. Efficient transformation of rice ( Oryza sativa L.) mediated by agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J, 1994: 271-282. |
| [21] | Liu W, Xie X, Ma X, Li J, Chen J, Liu Y. DSDecode: a web- based tool for decoding of sequencing chromatograms for genotyping of targeted mutations. Mol Plant, 2015,8:1431-1433. |
| [22] | Jackson D. No sex please, we’re (in) breeding. EMBO J, 2017,36:703-704. |
| [23] | 刘晨旭. 玉米单倍体诱导关键基因定位与克隆及新型高油诱导系选育研究. 中国农业大学博士学位论文, 北京, 2017. |
| Liu C X. Find Mapping and Cloning of Key QTLs Related to in vivo Haploid Induction and Super High Oil Haploid Inducer. PhD Dissertation of China Agricultural University, Beijing, China, 2017 (in Chinese with English abstract). | |
| [24] | Tian X L, Qin Y X, Chen B J, Liu C X, Wang L L, Li X L, Dong X, Liu L W, Chen S J. Hetero-fertilization together with failed egg-sperm cell fusion supports single fertilization involved in in vivo haploid induction in maize. J Exp Bot, 2018,69:4689-4701. |
| [1] | 田甜, 陈丽娟, 何华勤. 基于Meta-QTL和RNA-seq的整合分析挖掘水稻抗稻瘟病候选基因[J]. 作物学报, 2022, 48(6): 1372-1388. |
| [2] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [3] | 周文期, 强晓霞, 王森, 江静雯, 卫万荣. 水稻OsLPL2/PIR基因抗旱耐盐机制研究[J]. 作物学报, 2022, 48(6): 1401-1415. |
| [4] | 郑小龙, 周菁清, 白杨, 邵雅芳, 章林平, 胡培松, 魏祥进. 粳稻不同穗部籽粒的淀粉与垩白品质差异及分子机制[J]. 作物学报, 2022, 48(6): 1425-1436. |
| [5] | 颜佳倩, 顾逸彪, 薛张逸, 周天阳, 葛芊芊, 张耗, 刘立军, 王志琴, 顾骏飞, 杨建昌, 周振玲, 徐大勇. 耐盐性不同水稻品种对盐胁迫的响应差异及其机制[J]. 作物学报, 2022, 48(6): 1463-1475. |
| [6] | 杨建昌, 李超卿, 江贻. 稻米氨基酸含量和组分及其调控[J]. 作物学报, 2022, 48(5): 1037-1050. |
| [7] | 杨德卫, 王勋, 郑星星, 项信权, 崔海涛, 李生平, 唐定中. OsSAMS1在水稻稻瘟病抗性中的功能研究[J]. 作物学报, 2022, 48(5): 1119-1128. |
| [8] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [9] | 王小雷, 李炜星, 欧阳林娟, 徐杰, 陈小荣, 边建民, 胡丽芳, 彭小松, 贺晓鹏, 傅军如, 周大虎, 贺浩华, 孙晓棠, 朱昌兰. 基于染色体片段置换系群体检测水稻株型性状QTL[J]. 作物学报, 2022, 48(5): 1141-1151. |
| [10] | 王泽, 周钦阳, 刘聪, 穆悦, 郭威, 丁艳锋, 二宫正士. 基于无人机和地面图像的田间水稻冠层参数估测与评价[J]. 作物学报, 2022, 48(5): 1248-1261. |
| [11] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [12] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [13] | 杜晓芬, 王智兰, 韩康妮, 连世超, 李禹欣, 张林义, 王军. 谷子叶绿体基因RNA编辑位点的鉴定与分析[J]. 作物学报, 2022, 48(4): 873-885. |
| [14] | 王吕, 崔月贞, 吴玉红, 郝兴顺, 张春辉, 王俊义, 刘怡欣, 李小刚, 秦宇航. 绿肥稻秆协同还田下氮肥减量的增产和培肥短期效应[J]. 作物学报, 2022, 48(4): 952-961. |
| [15] | 巫燕飞, 胡琴, 周棋, 杜雪竹, 盛锋. 水稻延伸因子复合体家族基因鉴定及非生物胁迫诱导表达模式分析[J]. 作物学报, 2022, 48(3): 644-655. |
|
||