欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (7): 1832-1842.doi: 10.3724/SP.J.1006.2022.12028
黄福灯1( ), 黄妍2, 金泽艳2, 贺焕焕2, 李春寿1,*(), 程方民2, 潘刚2
), 黄妍2, 金泽艳2, 贺焕焕2, 李春寿1,*(), 程方民2, 潘刚2
HUANG Fu-Deng1(), HUANG Yan2, JIN Ze-Yan2, HE Huan-Huan2, LI Chun-Shou1,*(), CHENG Fang-Min2, PAN Gang2
摘要:
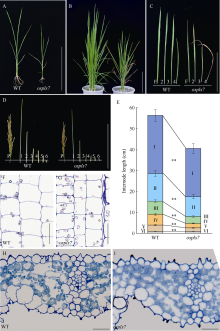
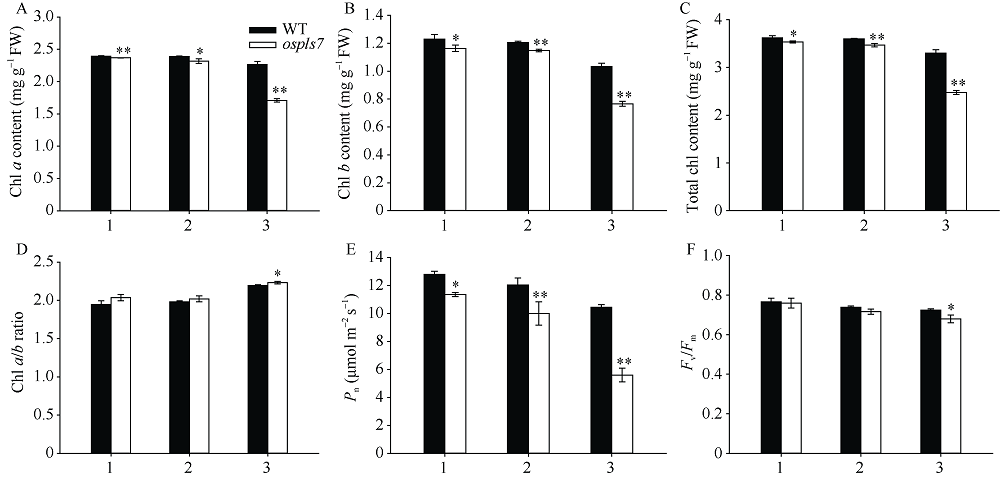
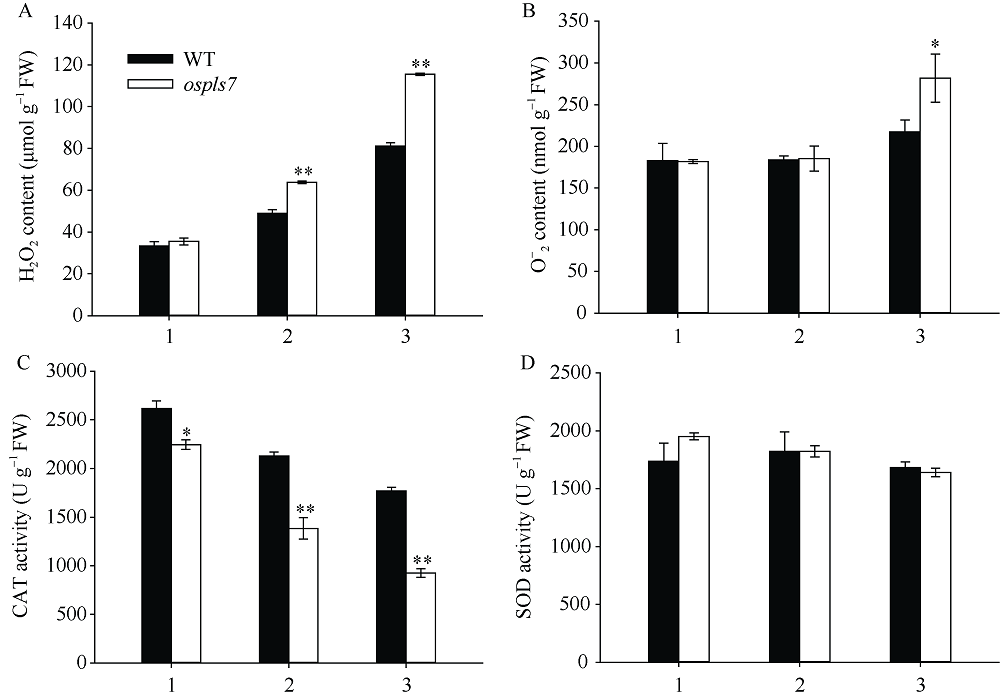
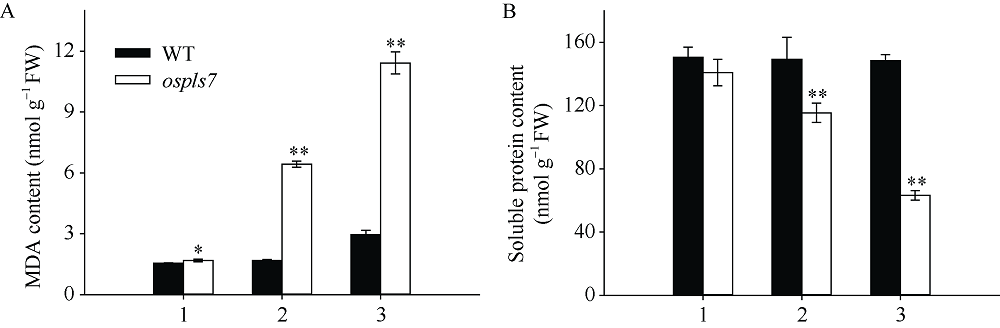
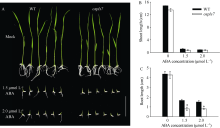
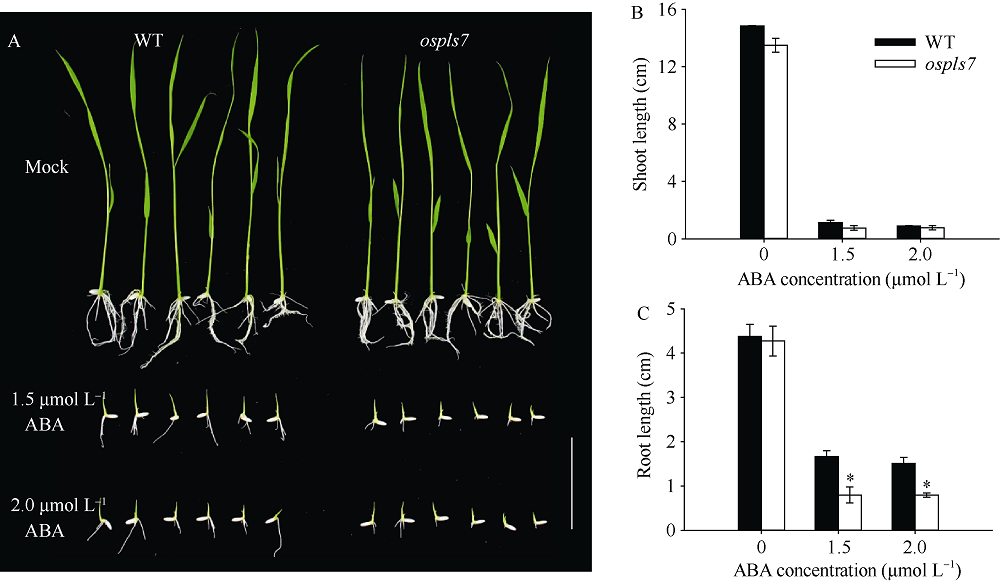
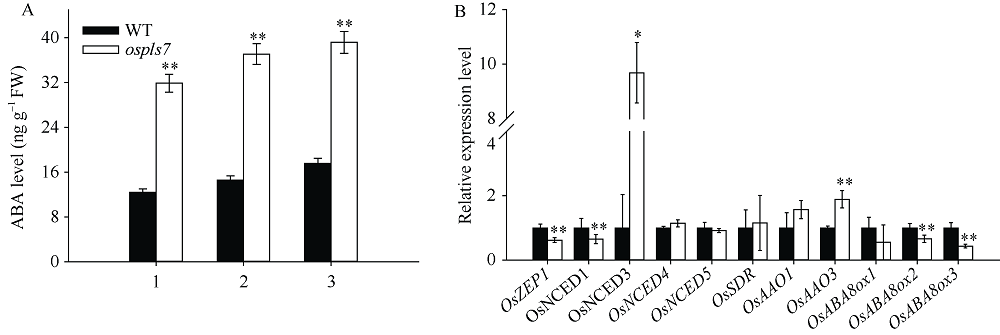
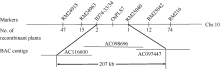
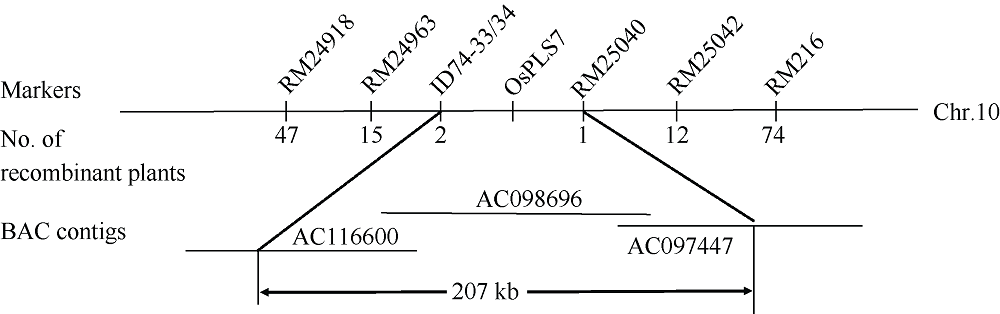
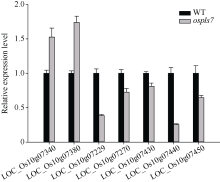
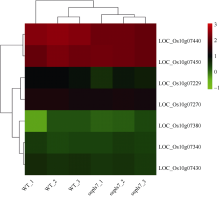
衰老是叶片发育的最后阶段, 但叶片, 尤其是功能叶, 早衰将影响作物的产量与品质, 因此, 研究叶片早衰的分子生理机制对培育耐早衰优良品种具有重要意义。通过60Co-γ辐射诱变旱稻Monolaya获得一个稳定遗传的叶片早衰突变体ospls7, 本文对其形态、叶片衰老生理特征、茎节细胞特性以及衰老性状遗传与基因定位等方面进行了研究。大田条件下, 突变体ospls7叶片早衰性状始于三至四叶期幼苗, 主要表现为: 叶尖及中上部叶边缘黄色褐化并最终枯萎, 成熟期穗长和各茎节长度均极显著短于野生型对照, 最终导致植株矮化。究其原因, 可能是由于突变体茎节细胞变短。叶片衰老生理结果表明, 与野生型对照相比, 孕穗期突变体ospls7倒二叶和倒三叶的叶绿素总含量、净光合速率、可溶性蛋白、过氧化氢酶活性均极显著降低, 致使其叶片中H2O2大量累积并引起丙二醛含量急剧增加。同时, 孕穗期突变体ospls7剑叶、倒二叶和倒三叶的内源ABA含量均极显著高于野生型对照, qRT-PCR结果证实ABA大量累积的原因在于ABA合成基因OsNCED3和OsAAO3显著上调, 而其代谢基因OsABA8ox2和OsABA8ox3则显著下调。遗传分析结果表明, 突变体ospls7的叶片早衰性状受单隐性核基因调控, 进一步利用图位克隆技术将该基因定位于10号染色体的InDel标记ID74-33/34和SSR标记RM25040之间, 物理距离为207 kb, 这些结果为最终克隆OsPLS7基因并研究其功能奠定了基础。
| [1] |
Lim P O, Kim H J, Nam H G. Leaf senescence. Annu Rev Plant Biol, 2007, 58: 115-136.
doi: 10.1146/annurev.arplant.57.032905.105316 |
| [2] |
Woo H R, Kim H J, Lim P O, Nam H G. Leaf senescence: systems and dynamics aspects. Annu Rev Plant Biol, 2019, 70: 347-376.
doi: 10.1146/annurev-arplant-050718-095859 |
| [3] | 田广丽, 孔亚丽, 张瑞卿, 周新国, 郭世伟. 不同氮水平下功能叶片数量和位置对水稻产量性状的影响. 植物营养与肥料学报, 2019, 25: 721-728. |
| Tian G L, Kong Y L, Zhang R Q, Zhou X G, Guo S W. Effects of the number and position of functional leaves on yield traits of rice under different nitrogen levels. Plant Nutr Fert Sci, 2019, 25: 721-728. (in Chinese with English abstract) | |
| [4] |
Thomas H, Smart C M. Crops that stay green. Ann Appl Biol, 1993, 123: 193-219.
doi: 10.1111/j.1744-7348.1993.tb04086.x |
| [5] | 陆定志, 潘裕才, 马跃芳, 林宗达, 鮑为群, 金逸民, 游树鹏. 杂交水稻抽穗结实期间叶片衰老的生理生化研究. 中国农业科学, 1988, 21(3): 21-26. |
| Lu D Z, Pan Y C, Ma Y F, Lin Z D, Bao W Q, Jin Y M, You S P. The physiological and biochemical research of leaf senescence during heading stage in hybrid rice. Sci Agric Sin, 1988, 21(3): 21-26. (in Chinese with English abstract) | |
| [6] | 刘道宏. 植物叶片的衰老. 植物生理学通讯, 1983, (2): 14-19. |
| Liu D H. Plant leaf senescence. Plant Physiol Commun, 1983, (2): 14-19. (in Chinese) | |
| [7] |
Leng Y J, Ye G Y, Zeng D L. Genetic dissection of leaf senescence in rice. Int J Mol Sci, 2017, 18: 2686.
doi: 10.3390/ijms18122686 |
| [8] |
Chen K, Guo J L, Ray A B, Song C P, Zhu J K, Zhao Y. Abscisic acid dynamics, signaling, and functions in plants. J Integr Plant Biol, 2020, 62: 25-54.
doi: 10.1111/jipb.12899 |
| [9] |
Asad M A U, Zakari S A, Zhao Q, Zhou L J, Ye Y, Cheng F M. Abiotic stresses intervene with ABA signaling to induce destructive metabolic pathways leading to death: premature leaf senescence in plants. Int J Mol Sci, 2019, 20: 256.
doi: 10.3390/ijms20020256 |
| [10] |
Mao C J, Lu S C, Lyu B, Zhang B, Shen J B, He J M, Luo L Q, Xi D D, Chen X, Ming F. A rice NAC transcription factor promotes leaf senescence via ABA biosynthesis. Plant Physiol, 2017, 174: 1747-1763.
doi: 10.1104/pp.17.00542 |
| [11] |
Sperotto R A, Ricachenevsky F K, Duarte G L, Boff T, Lopes K L, Sperb E R, Grusak M A, Fett J P. Identification of up-regulated genes in flag leaves during rice grain filling and characterization of OsNAC5, a new ABA-dependent transcription factor. Planta, 2009, 230: 985-1002.
doi: 10.1007/s00425-009-1000-9 pmid: 19697058 |
| [12] |
Liang C Z, Wang Y Q, Zhu Y N, Tang J Y, Hu B, Liu L C, Ou S J, Wu H K, Sun X H, Chu J F, Chu C C. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc Natl Acad Sci USA, 2014, 111: 10013-10018.
doi: 10.1073/pnas.1321568111 |
| [13] |
Kang K, Shim Y, Gi E, An G, Paek N C. Mutation of ONAC096 enhances grain yield by increasing panicle number and delaying leaf senescence during grain filling in rice. Int J Mol Sci, 2019, 20: 5241.
doi: 10.3390/ijms20205241 |
| [14] |
Sakuraba Y, Kim D, Han S H, Kim S H, Piao W L, Yanagisawa S, An G, Paek N C. Multilayered regulation of membrane-bound ONAC054 is essential for abscisic acid-induced leaf senescence in rice. Plant Cell, 2020, 32: 630-649.
doi: 10.1105/tpc.19.00569 |
| [15] |
Kim T, Kang K, Kim S H, An G, Paek N C. OsWRKY5 promotes rice leaf senescence via senescence-associated NAC and abscisic acid biosynthesis pathway. Int J Mol Sci, 2019, 20: 4437.
doi: 10.3390/ijms20184437 |
| [16] |
Piao W, Kim S H, Lee B D, An G, Sakuraba Y, Paek N C. Rice transcription factor OsMYB102 delays leaf senescence by down-regulating abscisic acid accumulation and signaling. J Exp Bot, 2019, 70: 2699-2715.
doi: 10.1093/jxb/erz095 |
| [17] |
Zhao Y, Chan Z L, Gao J H, Xing L, Cao M J, Yu C M, Hu Y L, You J, Shi H T, Zhu Y F, Gong Y H, Mu Z X, Wang H Q, Deng X, Wang P C, Bressan R A, Zhu J K. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc Natl Acad Sci USA, 2016, 113: 1949-1954.
doi: 10.1073/pnas.1522840113 |
| [18] |
Huang Y, Guo Y M, Liu Y T, Zhang F, Wang Z K, Wang H Y, Wang F, Li D P, Mao D D, Luan S, Liang M Z, Chen L B. 9-cis-epoxycarotenoid dioxygenase 3 regulates plant growth and enhances multi-abiotic stress tolerance in rice. Front Plant Sci, 2018, 9: 162.
doi: 10.3389/fpls.2018.00162 pmid: 29559982 |
| [19] |
Huang Y, Jiao Y, Xie N K, Guo Y M, Zhang F, Xiang Z P, Wang R, Wang F, Gao Q M, Tian L F, Li D P, Chen L B, Liang M Z. OsNCED5, a 9-cis-epoxycarotenoid dioxygenase gene, regulates salt and water stress tolerance and leaf senescence in rice. Plant Sci, 2019, 287: 110188.
doi: 10.1016/j.plantsci.2019.110188 |
| [20] |
He Y, Zhang Z H, Li L J, Tang S Q, Wu J L.Genetic and physio-biochemical characterization of a novel premature senescence leaf mutant in rice (Oryza sativa L.). Int J Mol Sci, 2018, 19: 2339.
doi: 10.3390/ijms19082339 |
| [21] |
Akhter D, Qin R, Nath U K, Alamin M, Jin X L, Shi C H. The brown midrib leaf (bml) mutation in rice (Oryza sativa L.) causes premature leaf senescence and the induction of defense responses. Genes (Basel), 2018, 9: 203.
doi: 10.3390/genes9040203 |
| [22] |
Wang S H, Lim J H, Kim S S, Cho S H, Yoo S C, Koh H J, Sakuraba Y, Paek N C. Mutation of SPOTTED LEAF3 (SPL3) impairs abscisic acid-responsive signaling and delays leaf senescence in rice. J Exp Bot, 2015, 66: 7045-7059.
doi: 10.1093/jxb/erv401 |
| [23] |
黄妍, 贺焕焕, 谢之耀, 李丹莹, 赵超越, 吴鑫, 黄福灯, 程方民, 潘刚. 水稻矮化宽叶突变体osdwl1的生理特性和基因定位. 作物学报, 2021, 47: 50-60.
doi: 10.3724/SP.J.1006.2021.92069 |
|
Huang Y, He H H, Xie Z Y, Li D Y, Zhao C Y, Wu X, Huang F D, Cheng F M, Pan G. Physiological characters and gene mapping of a dwarf and wide-leaf mutant osdwl1 in rice(Oryza sativa L.). Acta Agron Sin 2021, 47: 50-60 (in Chinese with English abstract).
doi: 10.3724/SP.J.1006.2021.92069 |
|
| [24] |
Chen W, Gong L, Guo Z L, Wang W S, Zhang H Y, Liu X Q, Yu S B, Xiong L Z, Luo J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: application in the study of rice metabolomics. Mol Plant, 2013, 6: 1769-1780.
doi: 10.1093/mp/sst080 pmid: 23702596 |
| [25] | 曹剑波, 程珂, 袁猛. 水稻组织半薄切片法. In: 袁猛, 都浩, 李香花编. Rice Protocol eBook. 袁猛, 都浩, 李香花编. Bio-101, 2018. https://bio-protocol.org/bio101/e1010142 . |
| Cao J B, Cheng K, Yuan M. Observation of rice tissue with semi-thin section. In: Yuan M, Du H, Li C H, eds. Rice Protocol eBook. Bio-101, 2018. https://bio-protocol.org/bio101/e1010142 . (in Chinese) | |
| [26] |
Murashige T, Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant, 1962, 15: 473-497.
doi: 10.1111/j.1399-3054.1962.tb08052.x |
| [27] |
Nakatsuka T, Nishihara M, Mishiba K, Yamamura S. Temporal expression of flavonoid biosynthesis-related genes regulates flower pigmentation in gentian plants. Plant Sci, 2005, 168: 1309-1318.
doi: 10.1016/j.plantsci.2005.01.009 |
| [28] |
Rogers S O, Bendich A J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol, 1985, 5: 69-76.
doi: 10.1007/BF00020088 pmid: 24306565 |
| [29] |
Jajic I, Sarna T, Strzalka K. Senescence, stress, and reactive oxygen species. Plants (Basel), 2015, 4: 393-411.
doi: 10.3390/plants4030393 |
| [30] |
Blokhina O, Virolainen E, Fagerstedt K V. Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot, 2003, 91: 179-194.
doi: 10.1093/aob/mcf118 |
| [31] |
张涛, 孙玉莹, 郑建敏, 程治军, 蒋开锋, 杨莉, 曹应江, 游书梅, 万建民, 郑家奎. 水稻早衰叶突变体PLS2的遗传分析与基因定位. 作物学报, 2014, 40: 2070-2080.
doi: 10.3724/SP.J.1006.2014.02070 |
|
Zhang T, Sun Y Y, Zheng J M, Cheng Z J, Jiang K F, Yang L, Cao Y J, You S M, Wan J M, Zheng J K. Genetic analysis and fine mapping of a premature leaf senescence mutant in rice (Orzya sativa L.). Acta Agron Sin, 2014, 40: 2070-2080. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2014.02070 |
|
| [32] | Rogers H, Munne-Bosch S. Production and scavenging of reactive oxygen species and redox signaling during leaf and flower senescence: similar but different. Plant Physiol, 2016, 17: 1560-1568. |
| [33] |
Breeze E, Harrison E, Mchattie S, Hughes L, Hickman R, Hill C, Kiddle S, Kim Y S, Penfold C A, Jenkins D, Zhang C, Morris K, Jenner C, Jackson S, Thomas B, Tabrett A, Legaie R, Moore J D, Wild D L, Ott S, Rand D, Beynon J, Denby K, Mead A, Buchanan-Wollaston V. High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant Cell, 2011, 23: 873-894.
doi: 10.1105/tpc.111.083345 |
| [34] |
Qi Y, Wang H, Zou Y, Liu C, Wang Y, Zhang W. Overexpression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett, 2011, 585: 231-239.
doi: 10.1016/j.febslet.2010.11.051 |
| [35] |
Kwon J, Mochida K, Wang K, Sekiguchi S, Sankai T, Aoki S, Ogura A, Yoshikawa Y, Wada K. Ubiquitin C-terminal hydrolase l-1 is essential for the early apoptotic wave of germinal cells and for sperm quality control during spermatogenesis. Biol Reprod, 2005, 73: 29-35.
doi: 10.1095/biolreprod.104.037077 |
| [36] |
Imin N, Kerim T, Weinman J J, Rolfe B G. Low temperature treatment at the young microspore stage induces protein changes in rice anthers. Mol Cell Proteom, 2006, 5: 274-292.
doi: 10.1074/mcp.M500242-MCP200 |
| [1] | 夏秀忠, 张宗琼, 杨行海, 荘洁, 曾宇, 邓国富, 宋国显, 黄欲晓, 农保选, 李丹婷. 广西水稻地方品种核心种质芽期耐盐性全基因组关联分析[J]. 作物学报, 2022, 48(8): 2007-2015. |
| [2] | 朱春权, 魏倩倩, 项兴佳, 胡文君, 徐青山, 曹小闯, 朱练峰, 孔亚丽, 刘佳, 金千瑜, 张均华. 褪黑素和茉莉酸甲酯基质育秧对水稻耐低温胁迫的调控作用[J]. 作物学报, 2022, 48(8): 2016-2027. |
| [3] | 刘昆, 黄健, 周沈琪, 张伟杨, 张耗, 顾骏飞, 刘立军, 杨建昌. 穗肥施氮量对不同穗型超级稻品种产量的影响及其机理[J]. 作物学报, 2022, 48(8): 2028-2040. |
| [4] | 委刚, 陈单阳, 任德勇, 杨宏霞, 伍靖雯, 冯萍, 王楠. 水稻细长秆突变体sr10的鉴定与基因定位[J]. 作物学报, 2022, 48(8): 2125-2133. |
| [5] | 周驰燕, 李国辉, 许轲, 张晨晖, 杨子君, 张芬芳, 霍中洋, 戴其根, 张洪程. 不同类型水稻品种茎叶维管束与同化物运转特征[J]. 作物学报, 2022, 48(8): 2053-2065. |
| [6] | 杜启迪, 郭会君, 熊宏春, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 宋希云, 刘录祥. 小麦顶端小穗退化突变体asd1基因定位[J]. 作物学报, 2022, 48(8): 1905-1913. |
| [7] | 陈驰, 陈代波, 孙志豪, 彭泽群, 贺登美, 张迎信, 程海涛, 于萍, 马兆慧, 宋建, 曹立勇, 程式华, 孙廉平, 占小登, 吕文彦. 水稻典败型隐性核雄性不育突变体ap90的鉴定与基因定位[J]. 作物学报, 2022, 48(7): 1569-1582. |
| [8] | 杨飞, 张征锋, 南波, 肖本泽. 水稻产量相关性状的全基因组关联分析及候选基因筛选[J]. 作物学报, 2022, 48(7): 1813-1821. |
| [9] | 田甜, 陈丽娟, 何华勤. 基于Meta-QTL和RNA-seq的整合分析挖掘水稻抗稻瘟病候选基因[J]. 作物学报, 2022, 48(6): 1372-1388. |
| [10] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [11] | 周文期, 强晓霞, 王森, 江静雯, 卫万荣. 水稻OsLPL2/PIR基因抗旱耐盐机制研究[J]. 作物学报, 2022, 48(6): 1401-1415. |
| [12] | 郑小龙, 周菁清, 白杨, 邵雅芳, 章林平, 胡培松, 魏祥进. 粳稻不同穗部籽粒的淀粉与垩白品质差异及分子机制[J]. 作物学报, 2022, 48(6): 1425-1436. |
| [13] | 颜佳倩, 顾逸彪, 薛张逸, 周天阳, 葛芊芊, 张耗, 刘立军, 王志琴, 顾骏飞, 杨建昌, 周振玲, 徐大勇. 耐盐性不同水稻品种对盐胁迫的响应差异及其机制[J]. 作物学报, 2022, 48(6): 1463-1475. |
| [14] | 杨建昌, 李超卿, 江贻. 稻米氨基酸含量和组分及其调控[J]. 作物学报, 2022, 48(5): 1037-1050. |
| [15] | 杨德卫, 王勋, 郑星星, 项信权, 崔海涛, 李生平, 唐定中. OsSAMS1在水稻稻瘟病抗性中的功能研究[J]. 作物学报, 2022, 48(5): 1119-1128. |
|